
Отличие прокариотической клетки от эукариотической кратко. Сравнительная характеристика строения клеток прокариот и эукариот
Прокариоты – древнейшие организмы, образующие самостоятельное царство. К прокариотам относятся бактерии, сине-зеленые «водоросли» и ряд других мелких групп.
Клетки прокариот не обладают, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов – линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли). Также к ним можно условно отнести постоянные внутриклеточные симбионты эукариотических клеток – митохондрии и пластиды.
Эукариоты (эвкариоты) (от греч. eu– хорошо, полностью иkaryon– ядро) – организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочечных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикрепленных изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты – митохондрии, а у водорослей и растений – также и пластиды.
2. Клетки эукариот. Строение и функции
К эукариотам относятся растения, животные, грибы.
Клеточной стенки у клеток животных нет. Она представлена голым протопластом. Пограничный слой клетки животных – гликокаликс – это верхний слой цитоплазматической мембраны, «усиленный» молекулами полисахаридов, которые входят в состав межклеточного вещества.
Митохондрии имеют складчатые кристы.
В клетках животных есть клеточный центр, состоящий из двух центриолей. Это говорит о том, что любая клетка животных потенциально способна к делению.
Включение в животной клетке представлено в виде зерен и капель (белки, жиры, углевод гликоген), конечных продуктов обмена, кристаллов солей, пигментов.
В клетках животных могут быть сократительные, пищеварительные, выделительные вакуоли небольших размеров.
В клетках нет пластид, включений в виде крахмальных зерен, крупных вакуолей, заполненных соком.
3. Сопоставление прокариотической и эукариотической клеток
Наиболее важным отличием эукариот от прокариот долгое время считалось наличие оформленного ядра и мембранных органоидов. Однако к 1970 – 1980-м гг. стало ясно, что это лишь следствие более глубинных различий в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у бактерий. (Таблица 16).
Именно наличие специфическим образом устроенного цитоскелета позволяет эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается, именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные симбионты, в том числе митохондрии и пластиды). Другая важнейшая функция цитоскелета эукариот – обеспечение деления ядра (митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических клеткок организовано проще). Различия в строении цитоскелета объясняют и другие отличия про- и эукариот. Например, постоянство и простоту форм прокариотических клеток и значительное разнообразие формы и способность к её изменению у эукариотических, а также относительно большие размеры последних.
Так, размеры прокариотических клеток составляют в среднем 0,5 – 5 мкм, размеры эукариотических – в среднем от 10 до 50 мкм. Кроме того, только среди эукариот попадаются поистине гигантские клетки, такие как массивные яйцеклетки акул или страусов (в птичьем яйце весь желток – это одна огромная яйцеклетка), нейроны крупных млекопитающих, отростки которых, укрепленные цитоскелетом, могут достигать десятков сантиметров в длину.
По своей структуре организмы могут одноклеточными и многоклеточными. Прокариоты преимущественно одноклеточны, за исключением некоторых цианобактерий и актиномицетов. Среди эукариот одноклеточное строение имеют простейшие, ряд грибов, некоторые водоросли. Все остальные формы многоклеточны. Считается, что одноклеточными были первые живые организмы Земли.
Самое очевидное отличие прокариот от эукариот заключается в наличии у последних ядра , что отражено в названии этих групп: «карио» с древнегреческого переводится как ядро, «про» - до, «эу» - хорошо. Отсюда прокариоты - это доядерные организмы, эукариоты - ядерные.
Однако это далеко не единственное и возможно не главное отличие прокариотических организмов от эукариот . В клетках прокариот вообще нет мембранных органоидов (за редким исключением) - митохондрий, хлоропластов, комплекса Гольджи, эндоплазматической сети, лизосом. Их функции выполняют выросты (впячивания) клеточной мембраны, на которых располагаются различные пигменты и ферменты, обеспечивающие процессы жизнедеятельности.
У прокариот нет характерных для эукариот хромосом. Их основной генетический материал - это нуклеоид , обычно имеющий форму кольца. В эукариотических клетках хромосомы представляют собой комплексы ДНК и белков-гистонов (играют важную роль в упаковке ДНК). Эти химические комплексы называются хроматином . Нуклеоид прокариот не содержит гистонов, а форму ему придают связанные с ним молекулы РНК.
Хромосомы эукариот находятся в ядре. У прокариот нуклеоид находится в цитоплазме и обычно крепится в одном месте к мембране клетки.
Кроме нуклеоида в прокариотических клетках бывает разное количество плазмид - нуклеоидов существенно меньшего размера, чем основной.
Количество генов в нуклеоиде прокариот на порядок меньше, чем в хромосомах. У эукариот есть множество генов, выполняющих регуляторную функцию по отношению к другим генам. Это дает возможность эукариотическим клеткам многоклеточного организма, содержащим одну и ту же генетическую информацию, специализироваться; изменяя свой метаболизм, более гибко реагировать на изменения внешней и внутренней среды. Отличается и структура генов. У прокариот гены в ДНК располагаются группами - оперонами. Каждый оперон транскрибируется как единое целое.
Отличия прокариот от эукариот есть и в процессах транскрипции и трансляции. Самое главное заключается в том, что в прокариотических клетках эти процессы могут протекать одновременно на одной молекуле матричной (информационной) РНК: в то время как она еще синтезируется на ДНК, на готовом ее конце уже «сидят» рибосомы и синтезируют белок. В эукариотических клетках мРНК после транскрипции претерпевает так называемое созревание. И только после этого на ней может синтезироваться белок.
Рибосомы прокариот меньше (коэффициент седиментации 70S), чем у эукариот (80S). Отличается количество белков и молекул РНК в составе субъединиц рибосом. Следует отметить, что рибосомы (а также генетический материал) митохондрий и хлоропластов схожи с прокариотами, что может говорить об их происхождении от древних прокариотических организмов, оказавшихся внутри клетки-хозяина.
Прокариоты отличаются обычно более сложным строением своих оболочек. Кроме цитоплазматической мембраны и клеточной стенки у них также имеется капсула и другие образования, в зависимости от типа прокариотического организма. Клеточная стенка выполняет опорную функцию и препятствует проникновению вредных веществ. В состав клеточной стенки бактерий входит муреин (гликопептид). Среди эукариот клеточная стенка есть у растений (ее основной компонент - целлюлоза), у грибов - хитин.
Прокариотические клетки делятся бинарным делением. У них нет сложных процессов клеточного деления (митоза и мейоза) , характерных для эукариот. Хотя перед делением нуклеоид удваивается, так же как хроматин в хромосомах. В жизненном цикле эукариот наблюдается чередование диплоидной и гаплоидной фаз. При этом обычно преобладает диплоидная фаза. В отличие от них у прокариот такого нет.
Клетки эукариот различны по размерам, но в любом случае существенно крупнее прокариотических (в десятки раз).
Питательные вещества в клетки прокариот поступают только с помощью осмоса. У эукариотических клеток кроме этого может также наблюдаться фаго- и пиноцитоз («захват» пищи и жидкости с помощью цитоплазматической мембраны).
В целом отличие прокариот от эукариот заключается в однозначно более сложном строении последних. Считается, что клетки прокариотического типа возникли путем абиогенеза (длительной химической эволюции в условиях ранней Земли). Эукариоты появились позже от прокариотов, путем их объединения (симбиотическая, а также химерная гипотезы) или эволюции отдельно взятых представителей (инвагинационная гипотеза). Сложность клеток эукариот позволила им организовать многоклеточный организм, в процессе эволюции обеспечить все основное разнообразие жизни на Земле.
Таблица отличий прокариот от эукариот
| Признак | Прокариоты | Эукариоты |
|---|---|---|
| Клеточное ядро | Нет | Есть |
| Мембранные органоиды | Нет. Их функции выполняют впячивания клеточной мембраны, на которых располагаются пигменты и ферменты. | Митохондрии, пластиды, лизосомы, ЭПС, комплекс Гольджи |
| Оболочки клетки | Более сложные, бывают различные капсулы. Клеточная стенка состоит из муреина. | Основной компонент клеточной стенки целлюлоза (у растений) или хитин (у грибов). У клеток животных клеточной стенки нет. |
| Генетический материал | Существенно меньше. Представлен нуклеоидом и плазмидами, которые меют кольцевую форму и находятся в цитоплазме. | Объем наследственной информации значительный. Хромосомы (состоят из ДНК и белков). Характерна диплоидность. |
| Деление | Бинарное деление клетки. | Есть митоз и мейоз. |
| Многоклеточность | Для прокариот не характерна. | Представлены как одноклеточными, так и многоклеточными формами. |
| Рибосомы | Мельче | Крупнее |
| Обмен веществ | Более разнообразный (гетеротрофы, фотосинтезирующие и хемосинтезирующие различными способами автотрофы; анаэробное и аэробное дыхание). | Автотрофность только у растений за счет фотосинтеза. Почти все эукариоты аэробы. |
| Происхождение | Из неживой природы в процессе химической и предбиологической эволюции. | От прокариот в процессе их биологической эволюции. |
Единство строения клеток.
Содержание любой клетки отделен от внешней среды особой структурой - плазматической мембраной (плазмалемма). Эта обособленность позволяет создавать внутри клетки совсем особая среда, не похоже на то, что его окружает. Поэтому в клетке могут происходить те процессы, которые не происходят нигде, их называют процессами жизнедеятельности.
Внутренняя среда живой клетки, ограниченное плазматической мембраной, называется цитоплазмой. Она включает гиалоплазму (основную прозрачную вещество) и клеточные органеллы, а также различные непостоянные структуры - включения. К органелл, которые есть в любой клетке, относятся также рибосомы, на которых происходит синтез белка.
Строение клеток эукариот.
Эукариоты - это организмы, клетки которых имеют ядро. Ядро - это самая органеллы эукариотической клетки, в которой хранится и из которой переписывается наследственная информация, записанная в хромосомах. Хромосома - это молекула ДНК, интегрированная с белками. В ядре содержится ядрышко - место, где образуются другие важные органеллы, участвующих в синтезе белка - рибосомы. Но рибосомы только формируются в ядре, а работают они (т.е. синтезируют белок) в цитоплазме. Часть из них находится в цитоплазме свободно, а часть прикрепляется к мембран, образуют сетку, которая получила название эндоплазматической.
Рибосомы - немембранни органеллы.
Эндоплазматическая сеть - это сеть канальцев, ограниченных мембранами. Существует два типа: гладкая и гранулярная. На мембранах гранулярной эндоплазматической сети расположены рибосомы, поэтому в ней происходит синтез и транспортировки белков. А гладкая эндоплазматическая сеть - это место синтеза и транспортировки углеводов и липидов. На ней рибосом нет.
Для синтеза белков, углеводов и жиров необходима энергия, которую в эукариотической клетке производят «энергетические станции» клетки - митохондрии.
Митохондрии - двомембранни органеллы, в которых осуществляется процесс клеточного дыхания. На мембранах митохондрий окисляются органические соединения и накапливается химическая энергия в виде особых энергетических молекул (АТФ).
В клетке также есть место, где органические соединения могут накапливаться и откуда они могут транспортироваться, - это аппарат Гольджи, система плоских мембранных мешочков. Он участвует в транспортировке белков, липидов, углеводов. В аппарате Гольджи образуются также органеллы внутриклеточного пищеварения - лизосомы.
Лизосомы - одномембранни органеллы, характерные для клеток животных, содержат ферменты, которые могут расщеплять белки, углеводы, нуклеиновые кислоты, липиды.
В клетке могут быть органеллы, не имеющие мембранной строения, например рибосомы и цитоскелет.
Цитоскелет - это опорно-двигательная система клетки, включает микрофиламенты, реснички, жгутики, клеточный центр, который производит микротрубочки и центриоли.
Существуют органеллы, характерные только для клеток растений, - пластиды. Бывают: хлоропласты, хромопласты и лейкопласты. В хлоропластах происходит процесс фотосинтеза.
В клетках растений также вакуоли - продукты жизнедеятельности клетки, являющиеся резервуарами воды и растворенных в ней соединений. В эукариотических организмов относятся растения, животные и грибы.
Строение клеток прокариот.
Прокариоты - одноклеточные организмы, в клетках которых нет ядра.
Прокариотические клетки малы по размерам, сохраняют генетический материал в форме кольцевой молекулы ДНК (нуклеоидом). В прокариотических организмов относятся бактерии и цианобактерии, которые раньше называли сине-зелеными водорослями.
Если в прокариот происходит процесс аэробного дыхания, то для этого используются специальные выпячивание плазматической мембраны - мезосомы. Если бактерии фотосинтезирующие, то процесс фотосинтеза происходит на фотосинтетических мембранах - тилакоидов.
Синтез белка в прокариот происходит на рибосомах. В прокариотических клетке мало органелл.
Гипотезы происхождения органелл эукариотических клеток.
Прокариотические клетки появились на Земле раньше, чем эукариотические.
1) симбиотические гипотеза объясняет механизм возникновения некоторых органоидов эукариотической клетки - митохондрий и фотосинтезирующих пластид.
2) Инвагинацыонная гипотеза - утверждает, что происхождение эукариотической клетки исходит из того, что предковой формы был аэробный прокариот. Органеллы в нем возникли в результате впячивания и отслоение частей оболочки с последующей функциональной специализацией в ядро, митохондрии, хлоропласты других органелл.
Строение эукариотической и прокариотической клеток. Эукариотическая клетка. Строение прокариотической клетки. Сравнение прокариотической и эукариотической клеток.
У современных и ископаемых организмов известны два типа клеток: прокариотическая и эукариотическая. Они столь резко различаются по особенностям строения, что это послужило для выделения двух надцарств живого мира - прокариот, т.е. доядерных, и эукариот, т.е. настоящих ядерных организмов. Промежуточные формы между этими крупнейшими таксонами живого пока неизвестны.
Основные признаки и отличия прокариотических и эукариотических клеток (таблица):
|
Признаки |
Прокариоты |
Эукариоты |
|
ЯДЕРНАЯ МЕМБРАНА |
Отсутствует |
Имеется |
|
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА |
Имеется |
Имеется |
|
МИТОХОНДРИИ |
Отсутствуют |
Имеются |
|
ЭПС |
Отсутствует |
Имеется |
|
РИБОСОМЫ |
Имеются |
Имеются |
|
ВАКУОЛИ |
Отсутствуют |
Имеются (особенно характерны для растений) |
|
ЛИЗОСОМЫ |
Отсутствуют |
Имеются |
|
КЛЕТОЧНАЯ СТЕНКА |
Имеется, состоит из сложного гетерополимерного вещества |
Отсутствует в животных клетках, в растительных состоит из целлюлозы |
|
КАПСУЛА |
Если имеется, то состоит из соединений белка и сахара |
Отсутствует |
|
КОМПЛЕКС ГОЛЬДЖИ |
Отсутствует |
Имеется |
|
ДЕЛЕНИЕ |
Простое |
Митоз, амитоз, мейоз |
Основное отличие прокариотических клеток от эукариотических заключается в том, что их ДНК не организована в хромосомы и не окружена ядерной оболочкой. Эукариотические клетки устроены значительно сложнее. Их ДНК, связанная с белком, организована в хромосомы, которые располагаются в особом образовании, по сути самом крупном органоиде клетки - ядре. Кроме того, внеядерное активное содержимое такой клетки разделено на отдельные отсеки с помощью эндоплазматической сети, образованной элементарной мембраной. Эукариотические клетки обычно крупнее прокариотических. Их размеры варьируют от 10 до 100 мкм, тогда как размеры клеток прокариот (различных бактерий, цианобактерий - сине- зеленых водорослей и некоторых других организмов), как правило, не превышают 10 мкм, часто составляя 2-3 мкм. В эукариотической клетке носители генов - хромосомы - находятся в морфологически оформленном ядре, отграниченном от остальной клетки мембраной. В исключительно тонких, прозрачных препаратах живые хромосомы можно видеть с помощью светового микроскопа. Чаще же их изучают на фиксированных и окрашенных препаратах.
Хромосомы состоят из ДНК, которая находится в комплексе с белками- гистонами, богатыми аминокислотами аргинином и лизином. Гистоны составляют значительную часть массы хромосом.
Эукариотическая клетка имеет разнообразные постоянные внутриклеточные структуры - органоиды (органеллы), отсутствующие в прокариотической клетке.
Прокариотические клетки могут делиться на равные части перетяжкой или почковаться, т.е. образовывать дочернюю клетку меньшего размера, чем материнская, но никогда не делятся путем митоза. Клетки эукариотических организмов, напротив, делятся путем митоза (исключая некоторые очень архаичные группы). Хромосомы при этом "расщепляются" продольно (точнее, каждая нить ДНК воспроизводит около себя свое подобие), и их "половинки" - хроматиды (полноценные копии нити ДНК) расходятся группами к противоположным полюсам клетки. Каждая из образующихся затем клеток получает одинаковый набор хромосом.
Рибосомы прокариотической клетки резко отличаются от рибосом эукариот по величине. Ряд процессов, свойственных цитоплазме многих эукариотических клеток, - фагоцитоз, пиноцитоз и циклоз (вращательное движение цитоплазмы) - у прокариот не обнаружен. Прокариотической клетке в процессе обмена веществ не требуется аскорбиновая кислота, но эукариотические не могут без нее обходиться.
Существенно различаются подвижные формы прокариотических и эукариотических клеток. Прокариоты имеют двигательные приспособления в виде жгутиков или ресничек, состоящих из белка флагеллина. Двигательные приспособления подвижных эукариотических клеток получили название ундулиподиев, закрепляющихся в клетке с помощью особых телец кинетосом. Электронная микроскопия выявила структурное сходство всех ундулиподиев эукариотических организмов и резкие их отличия от жгутиков прокариот
1. Строение эукариотической клетки.
Клетки, образующие ткани животных и растений, значительно различаются по форме, размерам и внутреннему строению. Однако все они обнаруживают сходство в главных чертах процессов жизнедеятельности, обмена веществ, в раздражимости, росте, развитии, способности к изменчивости.
Клетки всех типов содержат два основных компонента, тесно связанных между собой, — цитоплазму и ядро. Ядро отделено от цитоплазмы пористой мембраной и содержит ядерный сок, хроматин и ядрышко. Полужидкая цитоплазма заполняет всю клетку и пронизана многочисленными канальцами. Снаружи она покрыта цитоплазматической мембраной. В ней имеются специализированные структуры-органоиды,
присутствующие в клетке постоянно, и временные образования — включения. Мембранные органоиды
: наружная цитоплазматическая мембрана (HЦM), эндоплазматическая сеть (ЭПС), аппарат Гольджи, лизосомы, митохондрии и пластиды. В основе строения всех мембранных органоидов лежит биологическая мембрана. Все мембраны имеют принципиально единый план строения и состоят из двойного слоя фосфолипидов, в который с различных сторон ива разную глубину погружены белковые молекулы. Мембраны органоидов отличаются друг от друга лишь наборами входящих в них белков.
Цитоплазматическая мембрана.
У всех клеток растений, многоклеточных животных, у простейших и бактерий клеточная мембрана трехслойна: наружный и внутренний слои состоят из молекул белков, средний — из молекул липидов. Она ограничивает цитоплазму от внешней среды, окружает все органоиды клетки и представляет собой универсальную биологическую структуру. В некоторых клетках наружная оболочка образована несколькими мембранами, плотно прилегающими друг к другу. В таких случаях клеточная оболочка становится плотной и упругой и позволяет сохранить форму клетки, как, например, у эвглены и инфузории туфельки. У большинства растительных клеток, помимо мембраны, снаружи имеется еще толстая целлюлозная оболочка — клеточная стенка
. Она хорошо различима в обычном световом микроскопе и выполняет опорную функцию за счет жесткого наружного слоя, придающего клеткам четкую форму.
На поверхности клеток мембрана образует удлиненные выросты — микроворсинки, складки, впячивания и выпячивания, что во много раз увеличивает всасывающую или выделительную поверхность. С помощью мембранных выростов клетки соединяются друг с другом в тканях и органах многоклеточных организмов, на складках мембран располагаются разнообразные ферменты, участвующие в обмене веществ. Отграничивая клетку от окружающей среды, мембрана регулирует направление диффузии веществ и одновременно осуществляет активный перенос их внутрь клетки (накопление) или наружу (выделение). За счет этих свойств мембраны концентрация ионов калия, кальция, магния, фосфора в цитоплазме выше, а концентрация натрия и хлора ниже, чем в окружающей среде. Через поры наружной мембраны из внешней среды внутрь клетки проникают ионы, вода и мелкие молекулы других веществ. Проникновение в клетку относительно крупных твердых частиц осуществляется путем фагоцитоза
(от греч. "фаго” — пожираю, "питое” — клетка). При этом наружная мембрана в месте контакта с частицей прогибается внутрь клетки, увлекая частицу в глубь цитоплазмы, где она подвергается ферментативному расщеплению. Аналогичным путем в клетку попадают и капли жидких веществ; их поглощение называется пиноцитозом
(от греч. "пино” — пью, "цитос” — клетка). Наружная клеточная мембрана выполняет и другие важные биологические функции.
Цитоплазма
на 85 % состоит из воды, на 10 % — из белков, остальной объем приходится на долю липидов, углеводов, нуклеиновых кислот и минеральных соединений; все эти вещества образуют коллоидный раствор, близкий по консистенции глицерину. Коллоидное вещество клетки в зависимости от ее физиологического состояния и характера воздействия внешней среды имеет свойства и жидкости, и упругого, более плотного тела. Цитоплазма пронизана каналами различной формы и величины, которые получили название эндоплазматической сети.
Их стенки представляют собой мембраны, тесно контактирующие со всеми органоидами клетки и составляющие вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и перемещения веществ внутри клетки.
В стенках канальцев располагаются мельчайшие зернышки—гранулы, называемые рибосомами. Такая сеть канальцев называется гранулярной. Рибосомы могут располагаться на поверхности канальцев разрозненно или образуют комплексы из пяти-семи и более рибосом, называемые полисомами. Другие канальцы гранул не содержат, они составляют гладкую эндоплазматическую сеть. На стенках располагаются ферменты, участвующие в синтезе жиров и углеводов.
Внутренняя полость канальцев заполнена продуктами жизнедеятельности клетки. Внутриклеточные канальцы, образуя сложную ветвящуюся систему, регулируют перемещение и концентрацию веществ, разделяют различные молекулы органических веществ и этапы их, синтеза. На внутренней и внешней поверхности мембран, богатых ферментами, осуществляется синтез белков, жиров и углеводов, которые либо используются в обмене веществ, либо накапливаются в цитоплазме в качестве включений, либо выводятся наружу.
Рибосомы встречаются во всех типах клеток — от бактерий до клеток многоклеточных организмов. Это округлые тельца, состоящие из рибонуклеиновой кислоты (РНК) и белков почти в равном соотношении. В их состав непременно входит магний, присутствие которого поддерживает структуру рибосом. Рибосомы могут быть связаны с мембранами эндоплазматической сети, с наружной клеточной мембраной или свободно лежать в цитоплазме. В них осуществляется синтез белков. Рибосомы кроме цитоплазмы встречаются в ядре клетки. Они образуются в ядрышке и затем поступают в цитоплазму.
Комплекс Гольджи в растительных клетках имеет вид отдельных телец, окруженных мембранами. В животных клетках этот органоид представлен цистернами, канальцами и пузырьками. В мембранные трубки комплекса Гольджи из канальцев эндоплазматической сети поступают продукты секреции клетки, где они химически перестраиваются, уплотняются, а затем переходят в цитоплазму и либо используются самой клеткой, либо выводятся из нее. В цистернах комплекса Гольджи происходит синтез полисахаридов и их объединение с белками, в результате чего образуются гликопротеиды.
Митохондрии — небольшие тельца палочковидной формы, ограниченные двумя мембранами. От внутренней мембраны митохондрии отходят многочисленные складки — кристы, на их стенках располагаются разнообразные ферменты, с помощью которых осуществляется синтез высокоэнергетического вещества — аденозинтрифосфорной кислоты (АТФ). В зависимости от активности клетки и внешних воздействий митохондрии могут перемещаться, изменять свои размеры, форму. В митохондриях найдены рибосомы, фосфолипиды, РНК и ДНК. С присутствием ДНК в митохондриях связывают способность этих органоидов к размножению путем образования перетяжки или почкованием в период деления клетки, а также синтез части митохондриальных белков.
Лизосомы
- мелкие овальные образования, ограниченные мембраной и рассеянные по всей цитоплазме. Встречаются во всех клетках животных и растений. Они возникают в расширениях эндоплазматической сети и в комплексе Гольджи, здесь заполняются гидролитическими ферментами, а затем обособляются и поступают в цитоплазму. В обычных" условиях лизосомы переваривают частицы, попадающие в клетку путем фагоцитоза, и органоиды отмирающих клеток. Продукты лизиса выводятся через мембрану лизосомы в цитоплазму, где они включаются в состав новых молекул. При разрыве лизоеомной мембраны ферменты поступают в цитоплазму и переваривают ее содержимое, вызывая гибель клетки.
Пластиды
есть только в растительных клетках и встречаются, у большинства зеленых растений. В пластидах синтезируются и накапливаются органические вещества. Различают пластиды трех видов: хлоропласты, хромопласты и лейкопласты.
Хлоропласты —
зеленые пластиды, содержащие зеленый пигмент хлорофилл. Они находятся в листьях, молодых стеблях, незрелых плодах. Хлоропласты окружены двойной мембраной. У высших растений внутренняя часть хлоропластов заполнена полужидким веществом, в котором параллельно друг другу уложены пластинки. Парные мембраны пластинок, сливаясь, образуют стопки, содержащие хлорофилл. В каждой стопке хлоропластов высших растений чередуются слои молекул белка и молекул липидов, а между ними располагаются молекулы хлорофилла. Такая слоистая структура обеспечивает максимум свободных поверхностей и облегчает захват и перенос энергии в процессе фотосинтеза.
Хромопласты —
пластиды, в которых содержатся растительные пигменты (красный или бурый, желтый, оранжевый). Они сосредоточены в цитоплазме клеток цветков, стеблей, плодов, листьев растений и придают им соответствующую окраску. Хромопласты образуются из лейкопластов или хлоропластов в результате накопления пигментов каротиноидов.
Лейкопласты—бесцветные пластиды, располагающиеся в неокрашенных частях растений: в стеблях, корнях, луковицах и др. В лейкопластах одних клеток накапливаются зерна крахмала, в лейкопластах других клеток — масла, белки.
Все пластиды возникают из своих предшественников — пропластид. В них выявлена ДНК, которая контролирует размножение этих органоидов.
Клеточный центр,
или центросома, играет важную роль при делении, клетки и состоит из двух центриолей.
Он встречается у всех клеток животных и растений, кроме цветковых, низших грибов и некоторых, простейших. Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В делящейся клетке первым делится клеточный центр, одновременно образуется ахроматиновое веретено, ориентирующее хромосомы при расхождении их к полюсам. В дочерние клетки отходит по одной центриоле.
У многих растительных и животных клеток имеются органоиды специального назначения
: реснички,
выполняющие функцию движения (инфузории, клетки дыхательных путей), жгутики
(простейшие одноклеточные, мужские половые клетки у животных и растений и др.).
Включения - временные элемеаты, возникающие в клетке на определенной стадии ее жизнедеятельности в результате синтетической функции. Они либо используются, либо выводятся из клетки. Включениями являются также запасные питательные вещества: в растительных клетках—крахмал, капельки жира, белки, эфирные масла, многие органические кислоты, соли органических и неорганических кислот; в животных клетках - гликоген (в клетках печени и мышцах), капли жира (в подкожной клетчатке); Некоторые включения накапливаются в клетках как отбросы — в виде кристаллов, пигментов и др.
Вакуоли —
это полости, ограниченные мембраной; хорошо выражены в клетках растений и имеются у простейших. Возникают в разных участках расширений эндоплазматической сети. И постепенно отделяются от нее. Вакуоли поддерживают тургорное давление, в них сосредоточен клеточный или вакуолярный сок, молекулы которого определяют его осмотическую концентрацию. Считается, что первоначальные продукты синтеза - растворимые углеводы, белки, пектины и др. — накапливаются в цистернах эндоплазматической сети. Эти скопления и представляют собой зачатки будущих вакуолей.
Цитоскелет
.
Одной из отличительных особенностей эукариотической клетки является развитие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета тесно связаны с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме. Опорные элемеиты цитоплазмы определяют форму клетки, обеспечивают движение внутриклеточных структур и перемещение всей клетки.
Ядро
клетки играет основную роль в ее жизнедеятельности, с его удалением клетка прекращает свои функции и гибнет. В большинстве животных клеток одно ядро, но встречаются и многоядерные клетки (печень и мышцы человека, грибы, инфузории, зеленые водоросли). Эритроциты млекопитающих развиваются из клеток-предшественников, содержащих ядро, но зрелые эритроциты утрачивают его и живут недолго.
Ядро окружено двойной мембраной, пронизанной порами, посредством которых оно тесно связано с каналами эндоплазматической сети и цитоплазмой. Внутри ядра находится хроматин
— спирализованные участки хромосом. В период деления клетки они превращаются в палочковидные структуры, хорошо различимые в световой микроскоп. Хромосомы — это сложный комплекс белков с ДНК, называемый нуклеопротеидом.
Функции ядра состоят в регуляции всех жизненных отправлений клетки, которую оно осуществляет при помощи ДНК и РНК-материальных носителей наследственной информации. В ходе подготовки к делению клетки ДНК удваивается, в процессе митоза хромосомы расходятся и передаются дочерним клеткам, обеспечивая преемственность наследственной информации у каждого вида организмов.
Кариоплазма — жидкая фаза ядра, в которой в растворенном виде находятся продукты жизнедеятельности ядерных структур.
Ядрышко — обособленная, наиболее плотная часть ядра.
В состав ядрышка входят сложные белки и РНК, свободные или связанные фосфаты калия, магния, кальция, железа, цинка, а также рибосомы. Ядрышко исчезает перед началом деления клетки и вновь формируется в последней фазе деления.
Таким образом, клетка обладает тонкой и весьма сложной организацией. Обширная сеть цитоплазматических мембран и мембранный принцип строения органоидов позволяют разграничить множество одновременно протекающих в клетке химических реакций. Каждое из внутриклеточных образований имеет свою структуру и специфическую функцию, но только при их взаимодействии возможна гармоничная жизнедеятельность клетки.На основе такого взаимодействия вещества из окружающей среды поступают в клетку, а отработанные продукты выводятся из нее во внешнюю среду — так совершается обмен веществ. Совершенство структурной организации клетки могло возникнуть только в результате длительной биологической эволюции, в процессе которой выполняемые ею функции постепенно усложнялись.
Простейшие одноклеточные формы представляют собой и клетку, и организм со всеми его жизненными проявлениями. В многоклеточных организмах клетки образуют однородные группы — ткани. В свою очередь ткани формируют органы, системы, и их функции определяются общей жизнедеятельностью целостного организма.
2. Прокариотическая клетка.
К прокариотам относят бактерии и сине-зелёные водоросли (цианеи). Наследственный аппарат прокариот представлен одной кольцевой молекулой ДНК, не образующей связей с белками и содержащей по одной копии каждого гена — гаплоидные организмы. В цитоплазме имеется большое количество мелких рибосом; отсутствуют или слабо выражены внутренние мембраны. Ферменты пластического обмена расположены диффузно. Аппарат Гольджи представлен отдельными пузырьками. Ферментные системы энергетического обмена упорядоченно расположены на внутренней поверхности наружной цитоплазматической мембраны. Снаружи клетка окружена толстой клеточной стенкой. Многие прокариоты способны к спорообразованию в неблагоприятных условиях существования; при этом выделяется небольшой участок цитоплазмы содержащий ДНК, и окружается толстой многослойной капсулой. Процессы метаболизма внутри споры практически прекращаются. Попадая в благоприятные условия, спора преобразуется в активную клеточную форму. Размножение прокариот происходит простым делением надвое.
Средняя величина прокариотических клеток 5 мкм. У них нет никаких внутренних мембран, кроме впячиваний плазматической мембраны. Пласты отсутствуют. Вместо клеточного ядра имеется его эквивалент (нуклеоид), лишенный оболочки и состоящий из одной-единственной молекулы ДНК. Кроме того бактерии могут содержать ДНК в форме крошечных плазмид, сходных с внеядерными ДНК эукариот.
В прокариотических клетках, способных к фотосинтезу (сине-зеленые водоросли, зеленые и пурпурные бактерии) имеются различно структурированные крупные впячивания мембраны - тилакоиды, по своей функции соответствующие пластидам эукариот. Эти же тилакоиды или - в бесцветных клетках - более мелкие впячивания мембраны (а иногда даже сама плазматическая мембрана) в функциональном отношении заменяют митохондрии. Другие, сложно дифференцированные впячивания мембраны называют мезасомами; их функция не ясна.
Только некоторые органеллы прокариотической клетки гомологичны соответствующим органеллам эукариот. Для прокариот характерно наличие муреинового мешка - механически прочного элемента клеточной стенки
Сравнительная характеристика клеток растений, животных, бактерий, грибов
При сравнении бактерий с эукариотами можно выделить единственное сходство - наличие клеточной стенки, а вот сходства и различия эукариотических организмов заслуживают более пристального внимания. Следует начать сравнение с компонентов, которые свойственны и растениям, и животным, и грибам. Это ядро, митохондрии, Аппарат (комплекс) Гольджи, эндоплазматический ретикулум (или эндоплазматическая сеть) и лизосомы. Они характерны для всех организмов, имеют сходное строение и выполняют одинаковые функции. Теперь следует акцентировать внимание на различиях. Растительная клетка, в отличие от животной, имеет клеточную стенку, состоящую из целлюлозы. Кроме того, существую органеллы свойственные растительным клеткам - пластиды и вакуоли. Наличие этих компонентов обусловлено необходимостью растений поддерживать форму, при отсутствии скелета. Есть отличия и в особенностях роста. У растений он происходит в основном за счет увеличения размера вакуолей и растяжения клеток, в то время как у животных происходит увеличение объема цитоплазмы, а вакуоль вовсе отсутствует. Пластиды (хлоропласты, лейкопласты, хромопласты) характерны преимущественно для растений, поскольку их основная задача - это обеспечить автотрофный способ питания. У животных в противовес растениям существуют пищеварительные вакуоли, которые обеспечивают гетеротрофный способ питания. Грибы занимают особое положение и для их клеток характерны признаки свойственные и для растений, и для животных. Подобно животным грибам присущ гетеротрофный тип питания, содержащая хитин клеточная оболочка, а основным запасающим веществом является гликоген. В то же время для них, как для растений, характерен неограниченный рост, неспособность к передвижению и питание путем всасывания.
Сохранить в соцсетях:По строению клетки живые организмы делят на прокариот и эукариот . Клетки и тех и других окружены плазматической мембраной , снаружи от которой во многих случаях имеется клеточная стенка . Внутри клетки находится полужидкая цитоплазма . Однако клетки прокариот устроены значительно проще, чем клетки эукариот.
Основной генетический материал прокариот (от греч. про – до и карион – ядро) находится в цитоплазме в виде кольцевой молекулы ДНК. Эта молекула (нуклеоид ) не окружена ядерной оболочкой, характерной для эукариот, и прикрепляется к плазматической мембране (рис.1). Таким образом, прокариоты не имеют оформленного ядра. Кроме нуклеоида в прокариотической клетке часто встречается небольшая кольцевая молекула ДНК, называемая плазмидой . Плазмиды могут перемещаться из одной клетки в другую и встраиваться в основную молекулу ДНК.
Некоторые прокариоты имеют выросты плазматической мембраны: мезосомы, ламеллярные тилакоиды, хроматофоры . В них сосредоточены ферменты, участвующие в фотосинтезе и в процессах дыхания. Кроме того, мезосомы ассоциированы с синтезом ДНК и секрецией белка.
Клетки прокариот имеют небольшие размеры, их диаметр составляет 0,3–5 мкм. С наружной стороны плазматической мембраны всех прокариот (за исключением микоплазм) находится клеточная стенка . Она состоит из комплексов белков и олигосахаридов, уложенных слоями, защищает клетку и поддерживает ее форму. От плазматической мембраны она отделена небольшим межмембранным пространством.
В цитоплазме прокариот обнаруживаются только немембранные органоиды рибосомы . По структуре рибосомы прокариот и эукариот сходны, однако рибосомы прокариот имеют меньшие размеры и не прикрепляются к мембране, а располагаются прямо в цитоплазме.

Многие прокариоты подвижны и могут плавать или скользить с помощью жгутиков.
Размножаются прокариоты обычно путем деления надвое (бинарным ). Делению предшествует очень короткая стадия удвоения, или репликации, хромосом. Так что прокариоты – гаплоидные организмы.
К прокариотам относятся бактерии и синезеленые водоросли, или цианобактерии. Прокариоты появились на Земле около 3,5 млрд лет назад и были, вероятно, первой клеточной формой жизни, дав начало современным прокариотам и эукариотам.
Эукариоты (от греч. эу – истинный, карион – ядро) в отличие от прокариот, имеют оформленное ядро, окруженное ядерной оболочкой – двуслойной мембраной. Молекулы ДНК, обнаруживаемые в ядре, незамкнуты (линейные молекулы). Кроме ядра часть генетической информации содержится в ДНК митохондрий и хлоропластов. Эукариоты появились на Земле примерно 1,5 млрд лет назад.
В отличие от прокариот, представленных одиночными организмами и колониальными формами, эукариоты могут быть одноклеточными (например, амеба), колониальными (вольвокс) и многоклеточными организмами. Их делят на три больших царства: Животные, Растения и Грибы.
Диаметр клеток эукариот составляет 5–80 мкм. Как и прокариотические клетки, клетки эукариот окружены плазматической мембраной , состоящей из белков и липидов. Эта мембрана работает как селективный барьер, проницаемый для одних соединений и непроницаемый для других. Снаружи от плазматической мембраны расположена прочная клеточная стенка , которая у растений состоит главным образом из волокон целлюлозы, а у грибов – из хитина. Основная функция клеточной стенки – обеспечение постоянной формы клеток. Поскольку плазматическая мембрана проницаема для воды, а клетки растений и грибов обычно соприкасаются с растворами меньшей ионной силы, чем ионная сила раствора внутри клетки, вода будет поступать внутрь клеток. За счет этого объем клеток будет увеличиваться, плазматическая мембрана начнет растягиваться и может разорваться. Клеточная стенка препятствует увеличению объема и разрушению клетки.
У животных клеточная стенка отсутствует, но наружный слой плазматической мембраны обогащен углеводными компонентами. Этот наружный слой плазматической мембраны клеток животных называют гликокаликсом . Клетки многоклеточных животных не нуждаются в прочной клеточной стенке, поскольку есть другие механизмы, обеспечивающие регуляцию клеточного объема. Так как клетки многоклеточных животных и одноклеточные организмы, живущие в море, находятся в среде, в которой суммарная концентрация ионов близка к внутриклеточной концентрации ионов, клетки не набухают и не лопаются. Одноклеточные животные, живущие в пресной воде (амеба, инфузория туфелька), имеют сократительные вакуоли, которые постоянно выводят наружу поступающую внутрь клетки воду.
Структурные компоненты эукариотической клетки
Внутри клетки под плазматической мембраной находятся цитоплазма . Основное вещество цитоплазмы (гиалоплазма) представляет собой концентрированный раствор неорганических и органических соединений, главными компонентами которого являются белки. Это коллоидная система, которая может переходить из жидкого в гелеобразное состояние и обратно. Значительная часть белков цитоплазмы является ферментами, осуществляющими различные химические реакции. В гиалоплазме располагаются органоиды, выполняющие в клетке различные функции. Органоиды могут быть мембранными (ядро, аппарат Гольджи, эндоплазматический ретикулум, лизосомы, митохондрии, хлоропласты) и немембранными (клеточный центр, рибосомы, цитоскелет).
Мембранные органоидысновным компонентом мембранных органоидов является мембрана . Биологические мембраны построены по общему принципу, но химический состав мембран разных органоидов различен. Все клеточные мембраны – это тонкие пленки (толщиной 7–10 нм), основу которых составляет двойной слой липидов (бислой), расположенных так, что заряженные гидрофильные части молекул соприкасаются со средой, а гидрофобные остатки жирных кислот каждого монослоя направлены внутрь мембраны и соприкасаются друг с другом (рис. 3). В бислой липидов встроены молекулы белков (интегральные белки мембраны) таким образом, что гидрофобные части молекулы белка соприкасаются с жирнокислотными остатками молекул липидов, а гидрофильные части экспонированы в окружающую среду. Кроме этого часть растворимых (немембранных белков) соединяется с мембраной в основном за счет ионных взаимодействий (периферические белки мембраны). Ко многим белкам и липидам в составе мембран присоединены также углеводные фрагменты. Таким образом, биологические мембраны – это липидные пленки, в которые встроены интегральные белки.
Одна из основных функций мембран – создание границы между клеткой и окружающей средой и различными отсеками клетки. Липидный бислой проницаем в основном для жирорастворимых соединений и газов, гидрофильные вещества переносятся через мембраны с помощью специальных механизмов: низкомолекулярные – с помощью разнообразных переносчиков (каналов, насосов и др.), а высокомолекулярные – с помощью процессов экзо- и эндоцитоза (рис. 4).
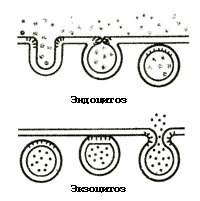
Рис. 4. Схема переноса веществ через мембрану
При эндоцитозе определенные вещества сорбируются на поверхности мембраны (за счет взаимодействия с белками мембраны). В этом месте образуется впячивание мембраны внутрь цитоплазмы. Затем от мембраны отделяется пузырек, внутри которого содержится переносимое соединение. Таким образом, эндоцитоз – это перенос в клетку высокомолекулярных соединений внешней среды, окруженных участком мембраны. Обратный процесс, то есть экзоцитоз – это перенос веществ из клетки наружу. Он происходит путем слияния с плазматической мембраной пузырька, заполненного транспортируемыми высокомолекулярными соединениями. Мембрана пузырька сливается с плазматической мембраной, а его содержимое изливается наружу.
Каналы, насосы и другие переносчики – это молекулы интегральных белков мембраны, обычно образующие в мембране пору.
Кроме функций разделения пространства и обеспечения избирательной проницаемости мембраны способны воспринимать сигналы. Эту функцию осуществляют белки-рецепторы, связывающие сигнальные молекулы. Отдельные белки мембраны являются ферментами, осуществляющими определенные химические реакции.
Ядро – крупный органоид клетки, окруженный ядерной оболочкой и имеющий обычно шаровидную форму. Ядро в клетке одно, и хотя встречаются многоядерные клетки (клетки скелетных мышц, некоторых грибов) или не имеющие ядра (эритроциты и тромбоциты млекопитающих), но эти клетки возникают из одноядерных клеток-предшественников.
Основная функция ядра – хранение, передача и реализация генетической информации . Здесь происходит удвоение молекул ДНК, в результате чего при делении дочерние клетки получают одинаковый генетический материал. В ядре с использованием в качестве матрицы отдельных участков молекул ДНК (генов) происходит синтез молекул РНК: информационных (иРНК), транспортных (тРНК) и рибосомальных (рРНК), необходимых для синтеза белка. В ядре осуществляется сборка субъединиц рибосом из молекул рРНК и белков, которые синтезируются в цитоплазме и переносятся в ядро.
Ядро состоит из ядерной оболочки, хроматина (хромосом), ядрышка и нуклеоплазмы (кариоплазмы).

Рис. 5. Структура хроматина: 1 – нуклеосома, 2 – ДНК
Под микроскопом внутри ядра видны зоны плотного вещества – хроматина. В неделящихся клетках он равномерно заполняет объем ядра или конденсируется в отдельных местах в виде более плотных участков и хорошо окрашивается основными красителями. Хроматин представляет собой комплекс ДНК и белков (рис. 5), большей частью положительно заряженных гистонов .
Количество молекул ДНК в ядре равно числу хромосом. Количество и форма хромосом являются уникальной характеристикой вида. В состав каждой из хромосом входит одна молекула ДНК, состоящая из двух связанных между собой нитей и имеющая вид двойной спирали толщиной 2 нм. Длина ее значительно превышает диаметр клетки: она может достигать нескольких сантиметров. Молекула ДНК заряжена отрицательно, поэтому сворачиваться (конденсироваться) она может только после связывания с положительно заряженными белками-гистонами (рис. 6).
Сначала двойная нить ДНК закручивается вокруг отдельных блоков гистонов, в каждый из которых входит 8 молекул белка, образуя структуру в виде «бусин на нитке» толщиной около 10 нм. Бусины называются нуклеосомами. В результате формирования нуклеосом длина молекулы ДНК уменьшается примерно в 7 раз. Далее нить с нуклеосомами сворачивается, формируя структуру в виде каната толщиной около 30 нм. Затем такой канат, изогнутый в виде петель, прикрепляется к белкам, образующим основу хромосомы. В результате образуется структура с толщиной около 300 нм. Дальнейшая конденсация этой структуры приводит к образованию хромосомы.
В период между делениями хромосома частично разворачивается. В результате этого отдельные участки молекулы ДНК, которые должны экспрессироваться в данной клетке, освобождаются от белков и вытягиваются, что делает возможным считывание с них информации путем синтеза молекул РНК.
Ядрышко – это тип матричной ДНК, отвечающей за синтез рРНК и собранной в отдельных участках ядра. Ядрышко – наиболее плотная структура ядра, оно не является отдельным органоидом, а представляет собой один из локусов хромосомы. В нем образуется рРНК, которая затем образует комплекс с белками, формируя субъединицы рибосом, которые уходят в цитоплазму.
Негистоновые белки ядра образуют внутри ядра структурную сеть. Она представлена слоем фибрилл, подстилающим ядерную оболочку. К ней прикрепляется внутриядерная сеть фибрилл, к которой присоединены фибриллы хроматина.
Ядерная оболочка состоит из двух мембран: внешней и внутренней, разделенных межмембранным пространством. Внешняя мембрана соприкасается с цитоплазмой, на ней могут находиться полирибосомы, а сама она может переходить в мембраны эндоплазматического ретикулума. Внутренняя мембрана связана с хроматином. Таким образом, ядерная оболочка обеспечивает фиксацию хромосомного материала в трехмерном пространстве ядра.
Оболочка ядра имеет круглые отверстия – ядерные поры (рис. 7). В области поры внешняя и внутренняя мембраны смыкаются и образуют отверстия, заполненные фибриллами и гранулами. Внутри поры располагается сложная система из белков, обеспечивающих избирательное связывание и перенос макромолекул. Количество ядерных пор зависит от интенсивности метаболизма клетки.
Эндоплазматический ретикулум , или эндоплазматическая сеть (ЭПР), представляет собой причудливую сеть каналов, вакуолей, уплощенных мешков, соединенных между собой и отделенных от гиалоплазмы мембраной (рис. 8).
Различают шероховатый и гладкий ЭПР. На мембранах шероховатого ЭПР находятся рибосомы (рис. 9), которые синтезируют белки, экскретируемые из клетки или встраивающиеся в плазматическую мембрану. Вновь синтезированный белок сходит с рибосомы и проходит через специальный канал внутрь полости эндоплазматического ретикулума, где он подвергается посттрансляционной модификации, например связыванию с углеводами, протеолитическому отщеплению части полипептидной цепи, образованию S–S-связей между остатками цистеина в цепи. Далее эти белки транспортируются в комплекс Гольджи, где входят либо в состав лизосом, либо секреторных гранул. В обоих случаях эти белки оказываются внутри мембранного пузырька (везикулы).

Рис. 9. Схема синтеза белка в
шероховатом ЭПР: 1 – малая и
2 – большая субъединицы рибосомы; 3 – молекула
рРНК;
4 – шероховатый ЭПР; 5 – вновь синтезируемый
белок
Гладкий ЭПР лишен рибосом. Его основная функция – синтез липидов и метаболизм углеводов. Он хорошо развит, например, в клетках коркового вещества надпочечников, где содержатся ферменты, обеспечивающие синтез стероидных гормонов. В гладком ЭПР в клетках печени находятся ферменты, осуществляющие окисление (детоксикацию) чужеродных для организма гидрофобных соединений, например лекарств.

Рис. 10. Аппарат Гольджи: 1 – пузырьки; 2 – цистерны
Комплекс Гольджи (рис. 10) состоит из 5–10 плоских ограниченных мембраной полостей, расположенных параллельно. Концевые части этих дискообразных структур имеют расширения. Таких образований в клетке может быть несколько. В зоне комплекса Гольджи находится большое количество мембранных пузырьков. Часть из них отшнуровывается от концевых частей основной структуры в виде секреторных гранул и лизосом. Часть мелких пузырьков (везикул), переносящих синтезированные в шероховатом ЭПР белки, перемещается к комплексу Гольджи и сливается с ним. Таким образом комплекс Гольджи участвует в накоплении и дальнейшей модификации продуктов, синтезированных в шероховатом ЭПР, и их сортировке.

Рис. 11. Образование и функции лизосом: 1 – фагосома; 2 – пиноцитозный пузырек; 3 – первичная лизосома; 4 – аппарат Гольджи; 5 – вторичная лизосома
Лизосомы – это вакуоли (рис. 11), ограниченные одной мембраной, которые отпочковываются от комплекса Гольджи. Внутри лизосом достаточно кислая среда (рН 4,9–5,2). Там располагаются гидролитические ферменты, расщепляющие различные полимеры при кислых рН (протеазы, нуклеазы, глюкозидазы, фосфатазы, липазы). Эти первичные лизосомы сливаются с эндоцитозными вакуолями, содержащими компоненты, которые должны расщепляться. Вещества, попавшие во вторичную лизосому, расщепляются до мономеров и переносятся через мембрану лизосомы в гиалоплазму. Таким образом, лизосомы участвуют в процессах внутриклеточного переваривания.
Митохондрии окружены двумя мембранами: наружной, отделяющей митохондрию от гиалоплазмы, и внутренней, отграничивающей ее внутреннее содержимое. Между ними располагается межмембранное пространство шириной 10–20 нм. Внутренняя мембрана образует многочисленные выросты (кристы ). В этой мембране располагаются ферменты, обеспечивающие окисление образовавшихся за пределами митохондрий аминокислот, сахаров, глицерина и жирных кислот (цикл Кребса) и осуществляющие перенос электронов в дыхательной цепи (схема). За счет переноса электронов по дыхательной цепи с высокого на более низкий энергетический уровень часть освобождающейся свободной энергии запасается в виде АТФ – универсальной энергетической валюты клетки. Таким образом, основная функция митохондрий – это окисление различных субстратов и синтез молекул АТФ.
Схема переноса двух электронов по дыхательной цепи
Внутри митохондрии находится кольцевая молекула ДНК, которая кодирует часть белков митохондрии. Во внутреннем пространстве митохондрий (матриксе) находятся рибосомы, похожие на рибосомы прокариот, которые и обеспечивают синтез этих белков.
Тот факт, что митохондрии имеют свою кольцевую ДНК и прокариотические рибосомы, привел к возникновению гипотезы, согласно которой митохондрия является потомком древней прокариотической клетки, когда-то попавшей внутрь эукариотической и в процессе эволюции взявшей на себя отдельные функции.
Рис. 12. Хлоропласты (А) и тилакоидные мембраны (Б)
Пластиды – органоиды растительной клетки, которые содержат пигменты. В хлоропластах содержится хлорофилл и каротиноиды, в хромопластах – каротиноиды, в лейкопластах пигментов нет. Пластиды окружены двойной мембраной. Внутри них располагается система мембран, имеющая форму плоских пузырьков, называемых тилакоидами (рис. 12). Тилакоиды уложены в стопки, напоминающие стопки тарелок. Пигменты встроены в мембраны тилакоидов. Их основная функция – поглощение света, энергия которого с помощью ферментов, встроенных в мембрану тилакоида, преобразуется в градиент ионов Н + на мембране тилакоида. Как и митохондрии, пластиды имеют собственную кольцевую ДНК и рибосомы прокариотического типа. По-видимому, пластиды также являются прокариотическим организмом, живущим в симбиозе с клетками эукариот.
Рибосомы –это немембранные клеточные органоиды, встречающиеся как в клетках про-, так и эукариот. Рибосомы эукариот больше по размеру, чем прокариотические, их размер составляет 25х20х20 нм. Состоит рибосома из большой и малой субъединиц, прилегающих друг к другу. Между субъединицами в функционирующей рибосоме располагается нить иРНК.
Каждая субъединица рибосомы построена из рРНК, плотно упакованной и связанной с белками. Рибосомы могут располагаться в цитоплазме свободно или быть связанными с мембранами ЭПР. Свободные рибосомы могут быть единичными, но могут образовывать полисомы, когда на одной нити иРНК располагается последовательно несколько рибосом. Основная функция рибосом – синтез белка.
Цитоскелет – это опорно-двигательная система клетки, включающая белковые нитчатые (фибриллярные) образования, являющиеся каркасом клетки и выполняющие двигательную функцию. Структуры цитоскелета динамичны, они возникают и распадаются. Цитоскелет представлен тремя типами образований: промежуточными филаментами (нити диаметром 10 нм), микрофиламент ы (нити диаметром 5–7 нм) и микротрубочками . Промежуточные филаменты – неветвящиеся белковые структуры в виде нитей, часто расположенные пучками. Их белковый состав различен в разных тканях: в эпителии они состоят из кератина, в фибробластах – из виментина, в мышечных клетках – из десмина. Промежуточные филаменты выполнят опорно-каркасную функцию.
Микрофиламенты – это фибриллярные структуры, расположенные непосредственно под плазматической мембраной в виде пучков или слоев. Они хорошо видны в ложноножках амебы, в движущихся отростках фибробластов, в микроворсинках кишечного эпителия (рис. 13). Микрофиламенты построены из сократительных белков актина и миозина и являются внутриклеточным сократительным аппаратом.
Микротрубочки входят в состав как временных, так и постоянных структур клетки. К временным относится веретено деления, элементы цитоскелета клеток между делениями, а к постоянным – реснички, жгутики и центриоли клеточного центра. Микротрубочки – это прямые полые цилиндры с диаметром около 24 нм, их стенки образованы округлыми молекулами белка тубулина. Под электронными микроскопом видно, что сечение микротрубочки образовано 13 субъединицами, соединенными в кольцо. Микротрубочки присутствуют в гиалоплазме всех эукариотических клеток. Одна из функций микротрубочек – создание каркаса внутри клеток. Кроме того, по микротрубочкам, как по рельсам, перемещаются мелкие везикулы.
Клеточный центр состоит из двух центриолей, расположенных под прямым углом друг к другу и связанных с ними микротрубочек. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления. Основой центриоли являются расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр, шириной 0,2 мкм и длиной 0,3–0,5 мкм. При подготовке клеток к делению центриоли расходятся и удваиваются. Перед митозом центриоли участвуют в образовании микротрубочек веретена деления. Клетки высших растений не имеют центриолей, но у них есть аналогичный центр организации микротрубочек.