
A sejt kémiai összetétele. Sejtszerkezet
ELMÉLET
A sejtszervecskék felépítése és funkciói
| Organoid név | A szerkezet jellemzői, funkciói |
| 1. Külső citoplazma membrán | Elválasztja a citoplazma tartalmát a külső környezettől; az ionok és a kis molekulák a pórusokon keresztül enzimek segítségével behatolhatnak a sejtbe; kommunikációt biztosít a sejtek között a szövetekben; A citoplazmatikus sejten kívül a növényi sejtnek cellulózból álló vastag membránja is van - egy sejtfal, amivel az állati sejtekben nincs. |
| 2. Citoplazma | A folyékony közeg, amelyben az organellumok és zárványok szuszpendálódnak, egy folyékony kolloid rendszerből áll, amelyben különféle anyagok molekulái vannak jelen. |
| 3. Plasztidok (leukoplasztok, kromoplasztok, kloroplasztok) | Csak növényi sejtekben található, két membránból álló organellumok. Zöld plasztiszok - speciális képződményekben klorofillt tartalmazó kloroplasztok - tilakoidok (granák), amelyekben fotoszintézis zajlik, képesek önmegújulásra (saját DNS-ük van) |
| 4. Endoplazmatikus retikulum | A sejtmag körül helyezkedik el, membránok, kiterjedt üregek és csatornák hálózata alkotja: sima Az EPS részt vesz a szén- és zsíranyagcserében; durva biztosítja a fehérjeszintézist a riboszómák segítségével |
| 5. Mitokondriumok | Két membrán szerkezet, a belső membránon kinövések - cristae - találhatók, amelyeken sok enzim található, az energiaanyagcsere oxigénszakaszának biztosítása(saját DNS-ük van) |
| 6. Vacuolák | A növényi sejt esszenciális szervei; oldott formában sok szerves anyagot, ásványi sót tartalmaz; állati sejtekben található |
| 7. Riboszómák | A gömb alakú részecskék, amelyek két alegységből állnak, szabadon helyezkednek el a citoplazmában, vagy az EPS membránjaihoz kapcsolódnak; fehérjeszintézist hajt végre |
| 8. Citoszkeleton | Mikrotubulusok és fehérjerostok kötegei, amelyek szorosan kapcsolódnak a külső membránhoz és a nukleáris burokhoz |
| 9. Flagella és csilló | A mozgásszervek általános szerkezeti tervvel rendelkeznek. A flagellák és a csillók mozgása az egyes párok mikrotubulusainak egymáshoz viszonyított elcsúszásának köszönhető. |
KÉRDÉSEK ÉS FELADATOK
- Mi a szénhidrátok funkciója a sejtben
1) katalitikus 2) energia 3) örökletes információ tárolása
4) részvétel a fehérje bioszintézisében
- Mi a DNS-molekulák funkciója egy sejtben?
1) épület 2) védő 3) örökletes információ hordozója
4) a napfény energia elnyelése
- A sejtben zajló bioszintézis során
1) szerves anyagok oxidációja 2) oxigénellátás és szén-dioxid eltávolítás
3) bonyolultabb szerves anyagok képződése, 4) keményítő glükózzá bomlása
- Az egyik rendelkezés sejtelmélet a dolog az
1) az élőlények sejtjei szerkezetükben és funkciójukban megegyeznek
2) a növényi szervezetek sejtekből állnak
3) az állati szervezetek sejtekből állnak
4) minden alacsonyabb és magasabb rendű szervezet sejtekből áll
- koncepció között riboszóma és fehérjeszintézis van egy bizonyos kapcsolat. Ugyanez a kapcsolat áll fenn a fogalom között sejt membrán és a következők egyike. Találd meg ezt a koncepciót.
1) anyagok szállítása 2) ATP szintézis 3) sejtosztódás 4) zsírszintézis
- A sejt belső környezetét ún
1) sejtmag 2) vakuólum 3) citoplazma 4) endoplazmatikus retikulum
- A sejt magjában található
1) lizoszómák 2) kromoszómák 3) plasztidok 4) mitokondriumok
- Milyen szerepet játszik a sejtmag a sejtben?
1) tápanyagellátást tartalmaz 2) kommunikál az organellumok között
3) elősegíti az anyagok bejutását a sejtbe 4) biztosítja az anyasejt és a lánya hasonlóságát
- A táplálékrészecskék emésztése és az elhalt sejtek eltávolítása a segítségével történik a szervezetben
1) Golgi-készülék 2) lizoszómák 3) riboszómák 4) endoplazmatikus retikulum
- Mi a riboszómák funkciója egy sejtben?
1) szénhidrátokat szintetizál, 2) fehérjeszintézist hajt végre
3) lebontja a fehérjéket aminosavakra 4) részt vesz a szervetlen anyagok felhalmozódásában
- A mitokondriumokban, a kloroplasztiszokkal ellentétben,
1) szénhidrátok szintézise 2) enzimek szintézise 3) ásványi anyagok oxidációja
4) szerves anyagok oxidációja
- A sejtekben hiányoznak a mitokondriumok
1) kakukk lenmoha 2) városi fecskék 3) papagájhalak 4) staphylococcus baktériumok
- A kloroplasztok a sejtekben találhatók
1) édesvízi hidra 2) fehér gomba micélium 3) éger szár 4) répalevél
- Az autotróf szervezetek sejtjei abban különböznek a heterotróf sejtektől, hogy vannak bennük
1) plasztidok 2) membránok 3) vakuolák 4) kromoszómák
- Sűrű héj, citoplazma, nukleáris anyag, riboszómák, plazmamembrán sejteket tartalmaz
1) algák 2) baktériumok 3) gombák 4) állatok
- Endoplazmatikus retikulum sejtben
1) szerves anyagok szállítását végzi
2) korlátozza a sejtet a környezettől vagy más sejtektől
3) részt vesz az energiaképzésben
4) megőrzi az örökletes információkat a sejt jeleiről és tulajdonságairól
- A gombasejtekben nem megy végbe a fotoszintézis, mert. hiányzik nekik
1) kromoszómák 2) riboszómák 3) mitokondriumok 4) plasztidok
- Nincs sejtszerkezetük, csak más élőlények sejtjeiben aktívak
1) baktériumok 2) vírusok 3) algák 4) protozoonok
- Az emberi és állati sejtekben energiaforrásként használják
1) hormonok és vitaminok 2) víz és szén-dioxid
3) szervetlen anyagok 4) fehérjék, zsírok és szénhidrátok
- A fogalomsorozatok közül melyik tükrözi a testet egyetlen rendszerként
1) Molekulák - sejtek - szövetek - szervek - szervrendszerek - szervezet
2) Szervrendszerek - szervek - szövetek - molekulák - sejtek - szervezet
3) Szerv - szövetek - szervezet - sejt - molekulák - szervrendszerek
4) Molekulák - szövetek - sejtek - szervek - szervrendszerek - szervezet
Sejt- egy élő rendszer elemi egysége. Az élő sejt különböző szerkezeteit, amelyek egy adott funkció ellátásáért felelősek, organellumoknak nevezzük, akárcsak az egész szervezet szerveit. A sejtben a specifikus funkciók az organellumok, bizonyos alakú intracelluláris struktúrák között oszlanak meg, mint például a sejtmag, a mitokondriumok stb.
Sejtszerkezetek:
Citoplazma. A sejt kötelező része, a plazmamembrán és a sejtmag közé zárva. Citoszol viszkózus vizes oldat különféle sókés szerves anyagok, amelyeket fehérjeszálak rendszere – citoszkeletonok – hatol át. A sejt kémiai és élettani folyamatainak nagy része a citoplazmában zajlik. Felépítése: citoszol, citoszkeleton. Funkciók: magában foglalja a különböző organellumokat, a sejt belső környezetét
plazma membrán. Az állatok, növények minden sejtjét a plazmamembrán korlátozza a környezettől vagy más sejtektől. Ennek a membránnak a vastagsága olyan kicsi (kb. 10 nm), hogy csak elektronmikroszkóppal látható.

Lipidek kettős réteget képeznek a membránban, és a fehérjék annak teljes vastagságában behatolnak, különböző mélységekbe merülnek a lipidrétegbe, vagy a membrán külső és belső felületén helyezkednek el. Az összes többi organellum membránjának szerkezete hasonló a plazmamembránhoz. Felépítése: kétrétegű lipidek, fehérjék, szénhidrátok. Funkciói: korlátozás, sejt alakjának megőrzése, károsodás elleni védelem, anyagok felvételének és eltávolításának szabályozója.
Lizoszómák. A lizoszómák hártyás organellumok. Ovális alakúak és 0,5 mikron átmérőjűek. Egy sor enzimet tartalmaznak, amelyek lebontják a szerves anyagokat. A lizoszómák membránja nagyon erős és megakadályozza saját enzimeinek behatolását a sejt citoplazmájába, de ha a lizoszómát bármilyen külső hatás károsítja, akkor az egész sejt vagy annak egy része elpusztul.
A lizoszómák a növények, állatok és gombák minden sejtjében megtalálhatók.
A különféle szerves részecskék emésztését végrehajtva a lizoszómák további "nyersanyagokat" biztosítanak a sejtben zajló kémiai és energiafolyamatokhoz. Az éhezés során a lizoszómasejtek megemésztenek bizonyos organellumokat anélkül, hogy megölnék a sejtet. Az ilyen részleges emésztés egy ideig biztosítja a sejt számára a szükséges minimális tápanyagot. Néha a lizoszómák egész sejteket és sejtcsoportokat emésztenek fel, ami alapvető szerepet játszik az állatok fejlődési folyamataiban. Példa erre a farok elvesztése az ebihal békává alakulása során. Szerkezete: ovális hólyagok, kívül membrán, belül enzimek. Funkciói: szerves anyagok lebontása, elhalt organellumok elpusztítása, elhasználódott sejtek elpusztítása.
Golgi komplexus. Az endoplazmatikus retikulum üregeinek és tubulusainak lumenébe jutó bioszintézis termékei koncentrálódnak és a Golgi-készülékben szállítódnak. Ennek az organellumnak a mérete 5-10 µm.

Szerkezet: membránokkal (vezikulák) körülvett üregek. Funkciói: felhalmozódás, csomagolás, szerves anyagok kiürítése, lizoszómák képzése
Endoplazmatikus retikulum. Az endoplazmatikus retikulum a szerves anyagok szintézisére és szállítására szolgáló rendszer a sejt citoplazmájában, amely összekapcsolt üregek áttört szerkezete.
Az endoplazmatikus retikulum membránjaihoz nagyszámú riboszóma kapcsolódik - a legkisebb sejtszervecskék, amelyek úgy néznek ki, mint egy 20 nm átmérőjű gömb. és RNS-ből és fehérjéből áll. A riboszómák azok, ahol a fehérjeszintézis zajlik. Ezután az újonnan szintetizált fehérjék bejutnak az üregek és tubulusok rendszerébe, amelyeken keresztül a sejt belsejében mozognak. Üregek, tubulusok, tubulusok membránokból, a riboszóma membránok felszínén. Funkciói: szerves anyagok szintézise riboszómák segítségével, anyagok szállítása.
Riboszómák. A riboszómák az endoplazmatikus retikulum membránjaihoz kapcsolódnak, vagy szabadon helyezkednek el a citoplazmában, csoportokba rendeződnek, és rajtuk szintetizálódnak a fehérjék. Fehérje összetétele, riboszómális RNS Funkciói: biztosítja a fehérje bioszintézist (fehérje molekula összeállítása ebből).
Mitokondriumok. A mitokondriumok energiaszervecskék. A mitokondriumok alakja eltérő, lehetnek a többi, rúd alakú, fonalas, átlagosan 1 mikron átmérőjűek. és 7 µm hosszú. A mitokondriumok száma a sejt funkcionális aktivitásától függ, és a rovarok repülő izmaiban elérheti a tízezret is. A mitokondriumokat kívülről egy külső membrán határolja, alatta egy belső membrán található, amely számos kinövést - cristae - képez.

A mitokondriumok belsejében RNS, DNS és riboszómák találhatók. Membránjaiba specifikus enzimek épülnek be, amelyek segítségével a táplálékanyagok energiája a mitokondriumokban ATP energiává alakul, amely a sejt és a szervezet egészének életéhez szükséges.
Membrán, mátrix, kinövések - cristae. Funkciói: ATP molekula szintézise, saját fehérjék, nukleinsavak, szénhidrátok, lipidek szintézise, saját riboszómák képzése.
plasztidok. Csak a növényi sejtben: leukoplasztok, kloroplasztok, kromoplasztok. Funkciói: tartalék szerves anyagok felhalmozása, beporzó rovarok vonzása, ATP és szénhidrátok szintézise. A kloroplasztok 4-6 mikron átmérőjű korong vagy golyó alakúak. Dupla membránnal - külső és belső. A kloroplaszton belül DNS-riboszómák és speciális membránszerkezetek - grana - találhatók, amelyek egymással és a kloroplaszt belső membránjával kapcsolódnak. Minden kloroplaszt körülbelül 50 szemcsét tartalmaz, a jobb fényelnyelés érdekében lépcsőzetesen. A klorofill a gran membránokban található, ennek köszönhetően a napfény energiája az ATP kémiai energiájává alakul. Az ATP energiáját a kloroplasztiszokban szerves vegyületek, elsősorban szénhidrátok szintézisére használják fel.
Kromoplasztok. A kromoplasztokban található vörös és sárga pigmentek adják a növény különböző részeinek vörös és sárga színét. sárgarépa, paradicsom gyümölcsök.
A leukoplasztok a tartalék tápanyag - a keményítő - felhalmozódásának helyei. Különösen sok leukoplaszt található a burgonyagumó sejtjeiben. Fényben a leukoplasztok kloroplasztokká alakulhatnak (aminek következtében a burgonyasejtek zöldre váltanak). Ősszel a kloroplasztiszok kromoplasztokká alakulnak, a zöld levelek és gyümölcsök pedig sárgák és pirosak.
Cell Center. Két hengerből, centriolokból áll, amelyek egymásra merőlegesen helyezkednek el. Funkciók: orsómenetek támogatása

A sejtzárványok vagy megjelennek a citoplazmában, vagy a sejt élete során eltűnnek.

A granulátum formájú, sűrű zárványok tartalék tápanyagokat (keményítő, fehérjék, cukrok, zsírok) vagy sejthulladékot tartalmaznak, amelyeket még nem lehet eltávolítani. A növényi sejtek minden plasztidja képes tartalék tápanyagokat szintetizálni és felhalmozni. A növényi sejtekben a tartalék tápanyagok felhalmozódása vakuólumokban történik.
Szemek, granulátumok, cseppek Funkciói: nem állandó képződmények, amelyek szerves anyagot és energiát tárolnak
Mag. Két membrán magburoka, maglé, nucleolus. Funkciói: örökletes információ tárolása a sejtben és szaporodása, RNS szintézis - információs, transzport, riboszómális. A spórák a magmembránban helyezkednek el, amelyen keresztül aktív anyagcsere megy végbe a sejtmag és a citoplazma között. A sejtmag nem csak az adott sejt összes jellemzőjéről és tulajdonságairól, a benne végbemenő folyamatokról (például fehérjeszintézis) tárol örökletes információkat, hanem a szervezet egészének jellemzőiről is. Az információkat a DNS-molekulák rögzítik, amelyek a kromoszómák fő részét képezik. A mag egy magot tartalmaz. A mag az örökletes információkat tartalmazó kromoszómák jelenléte miatt egy olyan központ funkcióit látja el, amely szabályozza a sejt minden létfontosságú tevékenységét és fejlődését.
Elmélet a biológia vizsga 4. feladatához
A sejt mint biológiai rendszer
A modern sejtelmélet, főbb rendelkezései, szerepe a modern természettudományos világkép kialakításában. A sejttel kapcsolatos ismeretek fejlesztése. Az élőlények sejtszerkezete a szerves világ egységének alapja, az élő természet kapcsolatának bizonyítéka
A modern sejtelmélet, főbb rendelkezései, szerepe a modern természettudományos világkép kialakításában
Az egyik alapfogalom a modern biológia az az elképzelés, hogy minden élő szervezetnek sejtszerkezete van. A sejt szerkezetének, létfontosságú tevékenységének és kölcsönhatásainak tanulmányozása környezet foglalkozik a tudománnyal citológia ma sejtbiológiának nevezik. A citológia megjelenését a sejtelmélet megfogalmazásának köszönheti (1838-1839, M. Schleiden, T. Schwann, kiegészítve 1855-ben R. Virchow-val).
sejtelmélet egy általános elképzelés a sejtek mint élő egységek felépítéséről és funkcióiról, szaporodásukról és a kialakulásában betöltött szerepéről többsejtű élőlények.
A sejtelmélet főbb rendelkezései:
- A sejt az élő szervezetek szerkezetének, élettevékenységének, növekedésének és fejlődésének egysége – a sejten kívül nincs élet.
- Sejt - egy rendszer, amely természetesen egymással összefüggő elemek halmazából áll, és egy bizonyos holisztikus formációt képvisel.
- Minden élőlény sejtje a maga módján hasonló. kémiai összetétel, szerkezete és funkciója.
- Új sejtek csak az anyasejtek osztódása („sejt a sejtből”) eredményeként jönnek létre.
- A többsejtű szervezetek sejtjei szöveteket alkotnak, a szervek pedig szövetekből épülnek fel. Egy szervezet egészének életét az alkotó sejtjeinek kölcsönhatása határozza meg.
- A többsejtű élőlények sejtjei teljes génkészlettel rendelkeznek, de abban különböznek egymástól, hogy különböző géncsoportok dolgoznak értük, ami a sejtek morfológiai és funkcionális sokféleségét - differenciálódását eredményezi.
A sejtelmélet megalkotásának köszönhetően világossá vált, hogy a sejt az élet legkisebb egysége, elemi élőrendszer, amely az élőlények minden jelével és tulajdonságával rendelkezik. A sejtelmélet megfogalmazása az öröklődésről és változékonyságról alkotott nézetek kialakulásának legfontosabb előfeltételévé vált, mivel természetük és eredendő mintáik azonosítása óhatatlanul az élő szervezetek szerkezetének egyetemességét sugallta. A sejtek kémiai összetételének és szerkezeti tervének egységének feltárása lendületet adott az élő szervezetek eredetével és fejlődésével kapcsolatos elképzelések kialakításához. Ezenkívül a többsejtű élőlények eredete egyetlen sejtből a folyamat során embrionális fejlődés a modern embriológia dogmájává vált.
A sejttel kapcsolatos ismeretek fejlesztése
A 17. századig az ember egyáltalán semmit sem tudott az őt körülvevő tárgyak mikroszerkezetéről, és szabad szemmel észlelte a világot. A mikrokozmosz tanulmányozására szolgáló műszert, a mikroszkópot hozzávetőleg 1590-ben találták fel G. és Z. Jansen holland mechanikusok, de tökéletlensége lehetetlenné tette kellően kicsi tárgyak vizsgálatát. Ezen a területen csak K. Drebbel (1572-1634) úgynevezett összetett mikroszkópjának megalkotása járult hozzá.
1665-ben R. Hooke (1635-1703) angol fizikus továbbfejlesztette a mikroszkóp kialakítását és a csiszolólencsék technológiáját, és a képminőség javulását akarva megvizsgálni az alatta lévő parafa metszeteket. faszénés élő növények. A metszeteken megtalálta a legkisebb méhsejtszerű pórusokat, és sejteknek nevezte őket (lat. cellula sejt, sejt). Érdekes megjegyezni, hogy R. Hooke a sejt fő alkotóelemének tekintette sejt membrán.
A 17. század második felében jelentek meg a legjelentősebb mikroszkóposok, M. Malpighi (1628-1694) és N. Gru (1641-1712) munkái, akik számos növény sejtszerkezetét is felfedezték.
Annak érdekében, hogy megbizonyosodjon arról, hogy amit R. Hooke és más tudósok láttak, az igaz, A. van Leeuwenhoek holland kereskedő, aki nem rendelkezett speciális végzettséggel, önállóan kifejlesztett egy mikroszkóp-tervet, amely alapvetően különbözött a meglévőtől, és javította a lencsegyártást. technológia. Ez lehetővé tette számára, hogy 275-300-szoros növekedést érjen el, és figyelembe vegye a szerkezet olyan részleteit, amelyek technikailag hozzáférhetetlenek voltak más tudósok számára. A. van Leeuwenhoek felülmúlhatatlan megfigyelő volt: mikroszkóp alatt gondosan felvázolta és leírta a látottakat, de nem igyekezett megmagyarázni. Egysejtű szervezeteket, köztük baktériumokat fedezett fel, sejtmagokat, kloroplasztokat, sejtfalvastagságokat talált a növényi sejtekben, de felfedezéseit sokkal később lehetett értékelni.
A 19. század első felében sorra következtek az élőlények belső szerkezetének összetevőinek felfedezései. G. Moth megkülönböztetett növényi sejtekben élő anyagés egy vizes folyadék - sejtnedv, talált pórusokat. R. Brown (1773-1858) angol botanikus 1831-ben fedezte fel a magot az orchideasejtekben, majd minden növényi sejtben megtalálták. J. Purkinje cseh tudós (1787-1869) bevezette a "protoplazma" (1840) kifejezést egy sejtmag nélküli sejt félig folyékony kocsonyás tartalmára. M. Schleiden (1804-1881) belga botanikus minden kortársánál tovább fejlődött, aki a magasabb rendű növények különböző sejtszerkezeteinek fejlődését és differenciálódását tanulmányozva bebizonyította, hogy minden növényi szervezet egy sejtből származik. A hagymapikkelysejtek magjában lekerekített nucleolus testeket is figyelembe vett (1842).
1827-ben K. Baer orosz embriológus felfedezte az emberek és más emlősök petéit, ezzel megcáfolva azt az elképzelést, hogy egy organizmus kizárólag hím ivarsejtekből fejlődik ki. Ezenkívül bebizonyította egy többsejtű állati szervezet kialakulását egyetlen sejtből - megtermékenyített tojásból, valamint a többsejtű állatok embrionális fejlődési szakaszainak hasonlóságát, ami eredetük egységét sugallta. A 19. század közepére felhalmozott információk általánosítást igényeltek, amely a sejtelméletté vált. A biológia megfogalmazását T. Schwann (1810-1882) német zoológusnak köszönheti, aki saját adatai és M. Schleiden növények fejlődésére vonatkozó következtetései alapján azt javasolta, hogy ha a mikroszkóp alatt látható bármely képződményben mag van jelen, akkor ez a képződmény sejt. E kritérium alapján fogalmazta meg T. Schwann a sejtelmélet főbb rendelkezéseit.
A német orvos és patológus, R. Virchow (1821-1902) egy másik fontos tételt is bevezetett ebbe az elméletbe: a sejtek csak az eredeti sejt osztódásával keletkeznek, vagyis a sejtek csak sejtekből ("cell from cell") keletkeznek.
A sejtelmélet megalkotása óta folyamatosan fejlődik a sejt mint a szervezet felépítésének, működésének és fejlődésének egysége doktrínája. A 19. század végére a mikroszkópos technika fejlődésének köszönhetően tisztázódott a sejt szerkezete, leírták az organellumokat - a sejt különböző funkciókat ellátó részeit, az új sejtek képződésének módszereit (mitózis, meiózis) tanulmányozták, és világossá vált a sejtszerkezetek kiemelkedő jelentősége az örökletes tulajdonságok átvitelében. A legújabb fizikai és kémiai kutatási módszerek alkalmazása lehetővé tette az örökletes információk tárolásának és továbbításának folyamataiba való elmélyülést, valamint az egyes sejtszerkezetek finomszerkezetének tanulmányozását. Mindez hozzájárult ahhoz, hogy a sejttudomány önálló tudásággá váljon - citológia.
Az élőlények sejtszerkezete, az összes szervezet sejtjeinek szerkezetének hasonlósága - a szerves világ egységének alapja, az élő természet kapcsolatának bizonyítéka
Minden jelenleg ismert élő szervezet (növények, állatok, gombák és baktériumok) sejtszerkezettel rendelkezik. Még a sejtszerkezettel nem rendelkező vírusok is csak sejtekben képesek szaporodni. A sejt az élővilág elemi szerkezeti és funkcionális egysége, amely minden megnyilvánulásában benne van, különösen az anyagcserében és az energiaátalakításban, a homeosztázisban, a növekedésben és fejlődésben, a szaporodásban és az ingerlékenységben. Ugyanakkor a sejtekben tárolják, dolgozzák fel és realizálják az örökletes információkat.
A sejtek sokfélesége ellenére a szerkezeti tervük ugyanaz: mindegyik tartalmaz örökletes apparátusbelemerülve citoplazmaés a környező cella plazma membrán.
A sejt hosszú evolúció eredményeként keletkezett szerves világ. A sejtek egyesítése többsejtű szervezetté nem egyszerű összegzés, hiszen minden egyes sejt, miközben megtartja az élő szervezetben rejlő összes jellemzőt, egyúttal új tulajdonságokra tesz szert egy bizonyos funkció ellátása miatt. Egyrészt egy többsejtű szervezet felosztható alkotórészeire - sejtekre, másrészt ezeket újra összerakva lehetetlen visszaállítani egy integrált szervezet funkcióit, hiszen új tulajdonságok csak a sejtek egymásra hatásában jelennek meg. a rendszer részei. Ebben nyilvánul meg az élőkre jellemző egyik fő minta, a diszkrét és az integrál egysége. A kis méret és a jelentős számú sejt nagy felületet hoz létre a többsejtű élőlényekben, ami a gyors anyagcsere biztosításához szükséges. Ezenkívül a test egy részének elhalása esetén a sejtek szaporodása miatt helyreállítható az integritása. A sejten kívül az öröklődő információk tárolása és továbbítása, az energia tárolása és átvitele, annak későbbi munkává alakítása lehetetlen. Végül egy többsejtű szervezetben a sejtek közötti funkciók megosztása bőséges lehetőséget biztosított az élőlények számára a környezetükhöz való alkalmazkodásra, és előfeltétele volt szerveződésük bonyolításának.
Így az összes élő szervezet sejtjei szerkezeti tervének egységének megállapítása bizonyítékul szolgált a Földön élő összes élet eredetének egységét illetően.
különféle sejtek. Prokarióta és eukarióta sejtek. Növények, állatok, baktériumok, gombák sejtjeinek összehasonlító jellemzői A sejtek sokfélesége
A sejtelmélet szerint a sejt az élőlények legkisebb szerkezeti és funkcionális egysége, amely az élőlények összes tulajdonságával rendelkezik. A sejtek száma szerint az organizmusokat egysejtűekre és többsejtűekre osztják. Az egysejtű szervezetek sejtjei független szervezetekként léteznek, és ellátják az élőlények összes funkcióját. Minden prokarióta és számos eukarióta (számos alga-, gomba- és protozoafaj) egysejtű, amely rendkívül változatos formájú és méretű lenyűgöző. A legtöbb élőlény azonban még mindig többsejtű. Sejtjeik bizonyos funkciók ellátására, szövetek és szervek kialakítására specializálódtak, ami nem csak morfológiai jellemzőkben tükröződik. Például az emberi test körülbelül 10 14 sejtből áll, amelyeket körülbelül 200 faj képvisel, amelyek alakja és mérete igen változatos.
A sejtek alakja lehet kerek, hengeres, köbös, prizmás, korong alakú, orsó alakú, csillag alakú stb. Tehát a tojások lekerekítettek, a hámsejtek hengeresek, köbösek és prizmásak, a vörösvértestek bikonkáv korong alakú, az izomszövet sejtjei orsó alakúak, a csillagok pedig az idegszövet sejtjei. Számos sejtnek egyáltalán nincs állandó alakja. Ezek közé tartoznak mindenekelőtt a vér leukocitái.
A sejtek mérete is jelentősen eltér: a többsejtű szervezet legtöbb sejtje 10 és 100 mikron közötti, a legkisebbé pedig 2-4 mikron. Az alsó határ abból adódik, hogy a sejtnek minimális anyag- és szerkezetkészlettel kell rendelkeznie az élet biztosításához, a túl nagy méretű sejtek pedig megakadályozzák az anyag- és energiacserét a környezettel, illetve a homeosztázis fenntartásának folyamatait is. Néhány sejt azonban szabad szemmel is látható. Mindenekelőtt ide tartoznak a görögdinnye és az almafák termésének sejtjei, valamint a halak és madarak tojásai. Még ha a cella egyik lineáris mérete meghaladja az átlagot, az összes többi megfelel a normának. Például egy neuron kinövés meghaladhatja az 1 métert, de átmérője továbbra is megfelel az átlagos értéknek. Nincs közvetlen kapcsolat a sejt mérete és a test mérete között. Tehát egy elefánt és egy egér izomsejtjei azonos méretűek.
Prokarióta és eukarióta sejtek
Mint fentebb említettük, a sejtek sok hasonló funkcionális tulajdonsággal és morfológiai jellemzővel rendelkeznek. Mindegyikük egy benne elmerült citoplazmából áll örökletes apparátus, és elkülönül a külső környezettől plazma membrán, vagy plasmalemma, amely nem zavarja az anyagcsere és az energia folyamatát. A membránon kívül a sejtnek különböző anyagokból álló sejtfala is lehet, amely a sejt védelmét szolgálja, egyfajta külső váza.
A citoplazma a sejt teljes tartalma, amely kitölti a plazmamembrán és a genetikai információkat tartalmazó szerkezet közötti teret. A fő anyagból áll - hialoplazma- és a benne elmerült organellumok és zárványok. Sejtszervecskék- ezek a sejt állandó komponensei, amelyek bizonyos funkciókat látnak el, a zárványok pedig a sejt élete során megjelenő és eltűnő komponensek, amelyek elsősorban raktározási vagy kiválasztó funkciókat látnak el. A zárványokat gyakran szilárdra és folyékonyra osztják. A szilárd zárványokat főleg szemcsék képviselik, és eltérő jellegűek is lehetnek, míg a vakuolák és zsírcseppek folyékony zárványnak minősülnek.

Jelenleg a sejtszerveződésnek két fő típusa van: prokarióta és eukarióta.
A prokarióta sejtnek nincs magja, genetikai információit nem választják el membránok a citoplazmától.
A citoplazma azon régióját, amely a prokarióta sejtben genetikai információkat tárol, ún nukleoid. A prokarióta sejtek citoplazmájában főként az organellumok egyik fajtája, a riboszómák találhatók, a membránokkal körülvett organellumok pedig teljesen hiányoznak. A baktériumok prokarióták.
Az eukarióta sejt olyan sejt, amelyben a fejlődés legalább egyik szakaszában van mag- egy speciális szerkezet, amelyben a DNS található.
Citoplazma eukarióta sejtek a membrán és nem membrán organellumok jelentős változatosságában különbözik. Nak nek eukarióta szervezetek növények, állatok és gombák. A prokarióta sejtek mérete általában egy nagyságrenddel kisebb, mint az eukarióta sejtek mérete. A legtöbb prokarióta egysejtű szervezet, míg az eukarióták többsejtűek.
Növények, állatok, baktériumok és gombák sejtjei szerkezetének összehasonlító jellemzői
A prokariótákra és eukariótákra jellemző tulajdonságokon kívül a növények, állatok, gombák és baktériumok sejtjei számos egyéb tulajdonsággal is rendelkeznek. Tehát a növényi sejtek specifikus organellumokat tartalmaznak - kloroplasztiszok, amelyek meghatározzák a fotoszintézis képességét, míg más szervezetekben ezek az organellumok nem találhatók meg. Természetesen ez nem jelenti azt, hogy más élőlények ne lennének képesek a fotoszintézisre, hiszen például a baktériumokban ez a plazmalemma és a citoplazmában lévő egyes membránvezikulák invaginációin fordul elő.
A növényi sejtek általában nagy, sejtnedvvel teli vakuolákat tartalmaznak. Az állatok, gombák és baktériumok sejtjeiben is megtalálhatók, de teljesen más eredetűek és más-más funkciót látnak el. A szilárd zárványok formájában található fő tartalékanyag a keményítő a növényekben, a glikogén állatokban és gombákban, valamint a glikogén vagy volutin a baktériumokban.
Ezen organizmuscsoportok másik megkülönböztető vonása a felszíni apparátus felépítése: az állati szervezetek sejtjei nem rendelkeznek sejtfallal, plazmamembránjukat csak vékony glikokalix borítja, míg az összes többinél van. Ez teljesen érthető, hiszen az állatok táplálkozási módja összefügg a fagocitózis folyamatában lévő táplálékrészecskék befogásával, és a sejtfal jelenléte megfosztaná őket ettől a lehetőségtől. A sejtfal részét képező anyag kémiai természete nem ugyanaz különféle csoportokélő szervezetek: ha növényekben cellulóz, akkor gombákban kitin, baktériumokban murein. Növények, állatok, gombák és baktériumok sejtszerkezetének összehasonlító jellemzői
| jel | baktériumok | Állatok | Gomba | Növények |
| Etetés módja | heterotróf vagy autotróf | Heterotróf | Heterotróf | autotróf |
| Örökletes információk szervezése | prokarióták | eukarióták | eukarióták | eukarióták |
| DNS lokalizáció | Nukleoid, plazmidok | sejtmag, mitokondrium | sejtmag, mitokondrium | Mag, mitokondriumok, plasztidok |
| plazma membrán | Van | Van | Van | Van |
| sejtfal | Mureinovaya | — | Kitinos | Cellulóz |
| Citoplazma | Van | Van | Van | Van |
| Sejtszervecskék | Riboszómák | Membrán és nem membrán, beleértve a sejtközpontot | Membrán és nem membrán | Membrán és nem membrán, beleértve a plasztidokat is |
| A mozgás szervei | Flagella és bolyhok | Flagella és csillók | Flagella és csillók | Flagella és csillók |
| Vacuolák | Ritkán | összehúzódó, emésztő | Néha | Központi vakuólum sejtnedvvel |
| Zárványok | Glikogén, volutin | glikogén | glikogén | Keményítő |
Az ábrán láthatók a vadon élő állatok különböző királyságai képviselőinek sejtszerkezetének különbségei.

A sejt kémiai összetétele. Makro- és mikroelemek. A sejtet alkotó szervetlen és szerves anyagok (fehérjék, nukleinsavak, szénhidrátok, lipidek, ATP) felépítésének és funkcióinak kapcsolata. A vegyszerek szerepe a sejtben és az emberi szervezetben
A sejt kémiai összetétele
Az élő szervezetek összetételében a D. I. Mengyelejev elemi periódusos rendszerének eddig felfedezett kémiai elemeinek többségét megtalálták. Egyrészt egyetlen olyan elemet sem tartalmaznak, amely az élettelen természetben ne lenne, másrészt koncentrációjuk az élettelen természetű testekben és az élő szervezetekben jelentősen eltér.
Ezek a kémiai elemek szervetlen és szerves anyagokat alkotnak. Annak ellenére, hogy az élő szervezetekben a szervetlen anyagok vannak túlsúlyban, a szerves anyagok határozzák meg kémiai összetételük egyediségét és általában az élet jelenségét, mivel ezeket főként az élőlények szintetizálják létfontosságú tevékenységük során, és fontos szerepet játszanak a szervezetben. reakciók.

A tudomány az élőlények kémiai összetételének és a bennük lezajló kémiai reakciók vizsgálatával foglalkozik. biokémia.
Meg kell jegyezni, hogy a vegyszerek tartalma a különböző sejtekben és szövetekben jelentősen eltérhet. Például míg az állati sejtekben a fehérjék dominálnak a szerves vegyületek között, addig a növényi sejtekben a szénhidrátok.
| Kémiai elem | földkéreg | Tengervíz | Élő organizmusok |
| O | 49.2 | 85.8 | 65-75 |
| C | 0.4 | 0.0035 | 15-18 |
| H | 1.0 | 10.67 | 8-10 |
| N | 0.04 | 0.37 | 1.5-3.0 |
| P | 0.1 | 0.003 | 0.20-1.0 |
| S | 0.15 | 0.09 | 0.15-0.2 |
| K | 2.35 | 0.04 | 0.15-0.4 |
| kb | 3.25 | 0.05 | 0.04-2.0 |
| Cl | 0.2 | 0.06 | 0.05-0.1 |
| mg | 2.35 | 0.14 | 0.02-0.03 |
| Na | 2.4 | 1.14 | 0.02-0.03 |
| Fe | 4.2 | 0.00015 | 0.01-0.015 |
| Zn | < 0.01 | 0.00015 | 0.0003 |
| Cu | < 0.01 | < 0.00001 | 0.0002 |
| én | < 0.01 | 0.000015 | 0.0001 |
| F | 0.1 | 2.07 | 0.0001 |
Makro- és mikroelemek
Körülbelül 80 kémiai elem található az élő szervezetekben, de ezek közül csak 27-nek van funkciója a sejtben és a szervezetben. A többi elem nyomokban jelen van, és úgy tűnik, hogy táplálékkal, vízzel és levegővel jut be. A szervezet kémiai elemeinek tartalma jelentősen változik. Koncentrációtól függően makroelemekre és mikroelemekre osztják őket.
Mindegyik koncentrációja makrotápanyagok a szervezetben meghaladja a 0,01%-ot, össztartalmuk pedig 99%. A makrotápanyagok közé tartozik az oxigén, a szén, a hidrogén, a nitrogén, a foszfor, a kén, a kálium, a kalcium, a nátrium, a klór, a magnézium és a vas. Ezen elemek közül az első négyet (oxigén, szén, hidrogén és nitrogén) is nevezik organogén, mivel a fő szerves vegyületek részét képezik. A foszfor és a kén számos szerves anyag, például fehérjék és nukleinsavak összetevői. A foszfor elengedhetetlen a csontok és a fogak kialakulásához.
A maradék makroelemek nélkül a szervezet normális működése lehetetlen. Tehát a kálium, a nátrium és a klór részt vesz a sejtek gerjesztési folyamataiban. A kálium számos enzim működéséhez és a víz sejtben való megtartásához is szükséges. A kalcium a növények sejtfalában, a csontokban, a fogakban és a puhatestű héjában található, és szükséges az izomösszehúzódáshoz és az intracelluláris mozgáshoz. A magnézium a klorofill összetevője - a pigment, amely biztosítja a fotoszintézis áramlását. Részt vesz a fehérje bioszintézisében is. A vas amellett, hogy a hemoglobin része, amely oxigént szállít a vérben, szükséges a légzési és fotoszintézis folyamataihoz, valamint számos enzim működéséhez.
nyomelemek 0,01%-nál kisebb koncentrációban vannak a szervezetben, és összkoncentrációjuk a sejtben még a 0,1%-ot sem éri el. A nyomelemek közé tartozik a cink, réz, mangán, kobalt, jód, fluor stb. A cink a hasnyálmirigy hormon inzulin molekulájának része, a réz a fotoszintézishez és a légzéshez szükséges. A kobalt a B12-vitamin összetevője, amelynek hiánya vérszegénységhez vezet. A jód szükséges a pajzsmirigyhormonok szintéziséhez, amelyek biztosítják az anyagcsere normális lefolyását, a fluor pedig a fogzománc kialakulásához kapcsolódik.
A makro- és mikroelemek anyagcseréjének hiánya és feleslege vagy megsértése egyaránt a fejlődéshez vezet különféle betegségek. Különösen a kalcium- és foszforhiány okoz angolkórt, a nitrogénhiány súlyos fehérjehiányt, a vashiány vérszegénységet, a jódhiány pedig a pajzsmirigyhormonok képződésének megsértését és az anyagcsere sebességének csökkenését okozza. A fluor bevitelének vízzel és táplálékkal való nagymértékű csökkentése a fogzománc megújulásának megsértését, és ennek következtében a fogszuvasodásra való hajlamot okozza. Az ólom szinte minden szervezetre mérgező. Feleslege visszafordíthatatlan agyi és központi idegrendszeri károsodást okoz, amely látás- és hallásvesztésben, álmatlanságban, veseelégtelenség, görcsök, valamint bénuláshoz és olyan betegségekhez is vezethet, mint a rák. Az akut ólommérgezést hirtelen hallucinációk kísérik, és kómával és halállal végződnek.
A makro- és mikroelemek hiánya az élelmiszerekben található tartalmuk növelésével és vizet inni valamint gyógyszerek szedésével. Tehát a jód megtalálható a tenger gyümölcseiben és a jódozott sóban, a kalcium a tojáshéjban stb.
A sejtet alkotó szervetlen és szerves anyagok (fehérjék, nukleinsavak, szénhidrátok, lipidek, ATP) felépítésének és funkcióinak kapcsolata. A vegyszerek szerepe a sejtben és az emberi szervezetben
szervetlen anyagok
A sejt kémiai elemei különféle - szervetlen és szerves - vegyületeket képeznek. A sejt szervetlen anyagai közé tartozik a víz, ásványi sók, savak stb., a szerves anyagok pedig a fehérjék, nukleinsavak, szénhidrátok, lipidek, ATP, vitaminok stb.
Víz(H 2 O) - a sejt leggyakoribb szervetlen anyaga, amely egyedülálló fizikai-kémiai tulajdonságokkal rendelkezik. Nincs íze, nincs színe, nincs szaga. Minden anyag sűrűségét és viszkozitását a víz becsüli meg. Sok más anyaghoz hasonlóan a víz is három halmazállapotú lehet: szilárd (jég), folyékony és gáznemű (gőz). A víz olvadáspontja $0°$C, a forráspontja $100°$C, azonban más anyagok vízben való oldódása megváltoztathatja ezeket a jellemzőket. A víz hőkapacitása is meglehetősen magas - 4200 kJ / mol K, ami lehetővé teszi, hogy részt vegyen a hőszabályozási folyamatokban. Egy vízmolekulában a hidrogénatomok 105°$-os szögben helyezkednek el, míg a közös elektronpárokat az elektronegatívabb oxigénatom húzza el. Ez határozza meg a vízmolekulák dipólus tulajdonságait (egyik vége pozitív, a másik negatív töltésű) és a vízmolekulák közötti hidrogénkötések kialakulásának lehetőségét. A vízmolekulák adhéziója a felületi feszültség, a kapilláris jelenség és a víz univerzális oldószer tulajdonságainak hátterében áll. Ennek eredményeként az összes anyagot vízben oldható (hidrofil) és abban oldhatatlan (hidrofób) anyagokra osztják. Ezeknek köszönhetően egyedi tulajdonságok Előre meghatározott, hogy a víz a földi élet alapja lett.

A szervezet sejtjeinek átlagos víztartalma nem azonos, és az életkorral változhat. Tehát egy másfél hónapos emberi embrióban a sejtek víztartalma eléri a 97,5% -ot, egy nyolc hónaposban - 83%, egy újszülöttben 74% -ra csökken, és egy felnőttben átlagosan 66%. A testsejtek azonban víztartalomban különböznek egymástól. Tehát a csontok körülbelül 20% vizet tartalmaznak, a máj - 70%, az agy pedig - 86%. Összességében elmondható a víz koncentrációja a sejtekben egyenesen arányos az anyagcsere sebességével.
ásványi sók lehet oldott vagy fel nem oldott állapotban. Oldható sók ionokra - kationokra és anionokra - disszociálnak. A legfontosabb kationok a kálium- és nátriumionok, amelyek elősegítik az anyagok membránon való átjutását, és részt vesznek az idegimpulzusok létrejöttében és vezetésében; valamint kalciumionok, amelyek részt vesznek az izomrostok összehúzódásában és a véralvadásban; magnézium, amely a klorofill része; vas, amely számos fehérje része, beleértve a hemoglobint is. A legfontosabb anionok a foszfát anion, amely az ATP és a nukleinsavak részét képezi, valamint a szénsavmaradék, amely tompítja a közeg pH-jának ingadozásait. Az ásványi sók ionjai biztosítják magának a víznek a sejtbe való bejutását és annak visszatartását is. Ha a sók koncentrációja a környezetben alacsonyabb, mint a sejtben, akkor a víz behatol a sejtbe. Az ionok meghatározzák a citoplazma puffertulajdonságait is, vagyis azt a képességét, hogy fenntartsa a citoplazma állandó enyhén lúgos pH-ját, annak ellenére, hogy a sejtben folyamatosan savas és lúgos termékek keletkeznek.
Oldhatatlan sók(CaCO 3, Ca 3 (PO 4) 2 stb.) az egysejtű és többsejtű állatok csontjainak, fogainak, héjának és héjának részei.
Ezenkívül más szervetlen vegyületek, például savak és oxidok is képződhetnek az élőlényekben. Tehát az emberi gyomor parietális sejtjei sósavat termelnek, amely aktiválja a pepszin emésztőenzimet, a szilícium-oxid pedig impregnálja a zsurló sejtfalát, és kovaalmahéjat képez. Az elmúlt években a nitrogén-monoxid (II) sejtekben és a szervezetben betöltött jelátvitelben betöltött szerepét is vizsgálták.
szerves anyag
A sejt szerves anyagainak általános jellemzői
A sejt szerves anyagait viszonylag egyszerű és összetettebb molekulák egyaránt képviselhetik. Azokban az esetekben, amikor egy összetett molekula (makromolekula) jelentős számú ismétlődő egyszerűbb molekulából jön létre, ún. polimerés szerkezeti egységek - monomerek. Attól függően, hogy a polimerek egységei ismétlődnek-e vagy sem, a következő kategóriába sorolhatók szabályos vagy szabálytalan. A polimerek a sejt szárazanyag-tömegének 90%-át teszik ki. A szerves vegyületek három fő osztályába tartoznak - szénhidrátok (poliszacharidok), fehérjék és nukleinsavak. A reguláris polimerek poliszacharidok, míg a fehérjék és a nukleinsavak szabálytalanok. A fehérjékben és a nukleinsavakban rendkívül fontos a monomerek sorrendje, mivel információs funkciót töltenek be.

Szénhidrát
Szénhidrát- ezek szerves vegyületek, amelyek főként három kémiai elemet tartalmaznak - szén, hidrogén és oxigén, bár számos szénhidrát tartalmaz nitrogént vagy ként is. A szénhidrátok általános képlete C m (H 2 O) n. Egyszerű és összetett szénhidrátokra oszthatók.
Egyszerű szénhidrátok (monoszacharidok) egyetlen cukormolekulát tartalmaznak, amelyet nem lehet egyszerűbbekre bontani. Ezek kristályos anyagok, édes ízűek és vízben jól oldódnak. A monoszacharidok aktívan részt vesznek a sejt anyagcseréjében, és az összetett szénhidrátok - oligoszacharidok és poliszacharidok - részét képezik.
A monoszacharidokat a szénatomok száma szerint osztályozzák (C3-C9), pl. pentózok(C 5) és hexózok(6-tól). A pentózok közé tartozik a ribóz és a dezoxiribóz. Ribóz az RNS és az ATP része. Dezoxiribóz a DNS összetevője. A hexózok (C 6 H 12 O 6) a glükóz, fruktóz, galaktóz stb. Szőlőcukor(szőlőcukor) minden szervezetben megtalálható, így az emberi vérben is, mivel energiatartalék. Számos összetett cukor része: szacharóz, laktóz, malátacukor, keményítő, cellulóz stb. Fruktóz(gyümölcscukor) a legnagyobb koncentrációban a gyümölcsökben, mézben, cukorrépa gyökérnövényekben található. Nemcsak az anyagcsere folyamatokban vesz részt aktívan, hanem része a szacharóznak és néhány poliszacharidnak is, mint például az inzulin.
A legtöbb monoszacharid képes ezüsttükörreakciót kiváltani és redukálni a rezet Fehling-folyadék (réz(II)-szulfát és kálium-nátrium-tartarát oldatának keveréke) hozzáadásával és forralással.

Nak nek oligoszacharidok magukban foglalják a több monoszacharidból képzett szénhidrátokat. Általában vízben is jól oldódnak és édes ízűek. E maradékok számától függően megkülönböztetünk diszacharidokat (két maradék), triszacharidokat (három) stb.. A diszacharidok közé tartozik a szacharóz, a laktóz, a maltóz stb. szacharóz(répa- vagy nádcukor) glükóz és fruktóz maradványokból áll, egyes növények raktározó szerveiben található. Különösen sok a szacharóz a cukorrépa és a cukornád gyökereiben, ahol ipari úton nyerik. A szénhidrátok édességének mércéjeként szolgál. Laktóz, vagy tejcukor, glükóz és galaktóz maradványai alkotják, megtalálhatók az anyai és tehéntej. Malátacukor(malátacukor) két glükózmaradékból áll. A poliszacharidok lebomlása során keletkezik a növényi magvakban és a emésztőrendszer ember, sörgyártásban használják.

Poliszacharidok olyan biopolimerek, amelyek monomerjei mono- vagy diszacharid maradékok. A legtöbb poliszacharid vízben oldhatatlan, és cukrozatlan ízű. Ezek közé tartozik a keményítő, a glikogén, a cellulóz és a kitin. Keményítő- Ez egy fehér porszerű anyag, amelyet nem nedvesít meg a víz, hanem forró vízzel lefőzve szuszpenziót képez - paszta. A keményítő valójában két polimerből áll, a kevésbé elágazó amilózból és az elágazóbb amilopektinből (2.9. ábra). Mind az amilóz, mind az amilopektin monomerje a glükóz. A keményítő a növények fő tartalékanyaga, amely nagy mennyiségben halmozódik fel magokban, gyümölcsökben, gumókban, rizómákban és a növények egyéb tárolószerveiben. A keményítőre adott minőségi reakció a jóddal való reakció, amelyben a keményítő kékeslila színűvé válik.

glikogén(állati keményítő) az állatok és gombák tartalék poliszacharidja, amely emberben a legnagyobb mennyiségben az izmokban és a májban halmozódik fel. Vízben is oldhatatlan, és cukrozatlan ízű. A glikogén monomerje a glükóz. A keményítőmolekulákhoz képest a glikogénmolekulák még elágazóbbak.
Cellulóz, vagy cellulóz, - a növények fő referencia poliszacharidja. A cellulóz monomerje a glükóz. Az el nem ágazó cellulózmolekulák kötegeket alkotnak, amelyek a növények sejtfalának részét képezik. A cellulóz a fa alapja, felhasználják az építőiparban, textil, papír, alkohol és számos szerves anyag gyártásában. A cellulóz kémiailag inert, nem oldódik sem savakban, sem lúgokban. Az emberi emésztőrendszer enzimei sem bontják le, de a vastagbélben lévő baktériumok segítik az emésztést. Ezenkívül a rost serkenti a gyomor-bél traktus falainak összehúzódását, segítve annak munkáját.

Chitin egy poliszacharid, amelynek monomerje egy nitrogéntartalmú monoszacharid. A gombák és ízeltlábúak héjának sejtfalának része. Az emberi emésztőrendszerben szintén nincs enzim a kitin emésztésére, csak néhány baktérium rendelkezik vele.
A szénhidrátok funkciói. A szénhidrátok műanyag (építő), energia, raktározási és támogató funkciókat látnak el a sejtben. Ezek alkotják a növények és gombák sejtfalát. 1 g szénhidrát lebontásának energiaértéke 17,2 kJ. A glükóz, a fruktóz, a szacharóz, a keményítő és a glikogén tartalék anyagok. A szénhidrátok részei lehetnek a komplex lipideknek és fehérjéknek is, amelyek glikolipideket és glikoproteineket képeznek, különösen a sejtmembránokban. Nem kevésbé fontos a szénhidrátok szerepe a környezeti jelek intercelluláris felismerésében és észlelésében, mivel receptorként működnek a glikoproteinek összetételében.
Lipidek
Lipidek A kis molekulatömegű, hidrofób tulajdonságokkal rendelkező anyagok kémiailag heterogén csoportja. Ezek az anyagok vízben nem oldódnak, emulziókat képeznek benne, de szerves oldószerekben jól oldódnak. A lipidek olajos tapintásúak, sokuk jellegzetes, nem száradó nyomokat hagy a papíron. A fehérjékkel és szénhidrátokkal együtt a sejtek egyik fő összetevője. A különböző sejtek lipidtartalma nem egyforma, különösen sok van belőlük egyes növények magjában, termésében, a májban, szívben, vérben.
A molekula szerkezetétől függően a lipideket egyszerű és összetett csoportokra osztják. Nak nek egyszerű a lipidek közé tartoznak a semleges lipidek (zsírok), viaszok és szteroidok. Összetett a lipidek egy másik, nem lipid komponenst is tartalmaznak. Ezek közül a legfontosabbak a foszfolipidek, glikolipidek stb.
Zsírok a háromértékű alkohol glicerin és a magasabb zsírsavak észterei. A legtöbb zsírsav 14-22 szénatomot tartalmaz. Ezek között vannak telített és telítetlenek, azaz kettős kötéseket tartalmazók. A telített zsírsavak közül a palmitinsav és a sztearinsav, a telítetlen zsírsavak közül pedig az olajsav a leggyakoribb. Egyes telítetlen zsírsavak az emberi szervezetben nem vagy nem megfelelő mennyiségben szintetizálódnak, ezért nélkülözhetetlenek. A glicerinmaradékok hidrofil fejeket, míg a zsírsavmaradékok hidrofób végeket képeznek.

A zsírok főként raktározási funkciót töltenek be a sejtekben, és energiaforrásként szolgálnak. Gazdagok a bőr alatti zsírszövetben, amely ütéselnyelő és hőszigetelő funkciót lát el, a vízi állatokban pedig a felhajtóerőt is növeli. A növényi zsírok többnyire telítetlen zsírsavakat tartalmaznak, ennek következtében folyékonyak és ún. olajok. Az olajok számos növény magjában találhatók, mint például a napraforgó, szójabab, repce stb.
Viaszok zsírsavak és zsíralkoholok észterei és keverékei. A növényekben a levél felületén filmréteget képeznek, amely megvédi a párolgást, a kórokozók behatolását stb. Számos állatnál befedik a testet, vagy lépek kialakítására szolgálnak.
Nak nek szteroidok közé tartoznak a lipidek, például a koleszterin, amely a sejtmembránok alapvető összetevője, valamint a nemi hormonok ösztradiol, tesztoszteron, D-vitamin stb.
Foszfolipidek, a glicerin és zsírsavak mellett ortofoszforsav maradékot is tartalmaz. A sejtmembránok részét képezik, és biztosítják védő tulajdonságaikat.
Glikolipidek membránok alkotóelemei is, de tartalmuk alacsony. A glikolipidek nem lipid része szénhidrátok.
A lipidek funkciói. A lipidek képlékeny (építő), energia, raktározó, védő, kiválasztó és szabályozó funkciókat látnak el a sejtben, emellett vitaminok. A sejtmembránok nélkülözhetetlen alkotóeleme. 1 g lipid felhasadásakor 38,9 kJ energia szabadul fel. A tartalékban lerakódnak a növények és állatok különféle szerveiben. Ezenkívül a bőr alatti zsírszövet véd belső szervek hipotermia vagy túlmelegedés, valamint sokk miatt. A lipidek szabályozó funkciója annak a ténynek köszönhető, hogy egy részük hormon. A rovarok zsírteste a kiürülést szolgálja.
Mókusok
Mókusok- Ezek nagy molekulatömegű vegyületek, biopolimerek, amelyek monomerjei peptidkötésekkel összekapcsolt aminosavak.
aminosav aminocsoporttal, karboxilcsoporttal és gyökkel rendelkező szerves vegyületnek nevezzük. Összesen mintegy 200 aminosav található a természetben, amelyek gyökökben és kölcsönös elrendeződésben különböznek egymástól. funkcionális csoportok, de csak 20 közülük lehet fehérjék része. Ezeket az aminosavakat ún proteinogén.

Sajnos nem minden proteinogén aminosav szintetizálható az emberi szervezetben, ezért felcserélhetők és pótolhatatlanok. Nem esszenciális aminosavak az emberi szervezetben a szükséges mennyiségben képződnek, és pótolhatatlan- Nem. Élelmiszerből kell származniuk, de részben a bél mikroorganizmusai is szintetizálhatják. 8 teljesen esszenciális aminosav van, ezek közé tartozik a valin, izoleucin, leucin, lizin, metionin, treonin, triptofán és fenilalanin. Annak ellenére, hogy abszolút az összes proteinogén aminosav szintetizálódik a növényekben, a növényi fehérjék hiányosak, mert nem tartalmazzák az aminosavak teljes készletét, ráadásul a növények vegetatív részeiben a fehérje jelenléte ritkán haladja meg a fehérje 1-2%-át. tömeg. Ezért nemcsak növényi, hanem állati eredetű fehérjéket is kell enni.
A peptidkötésekkel összekapcsolt két aminosavból álló szekvenciát nevezzük dipeptid, a háromból tripeptid stb. A peptidek között vannak olyan fontos vegyületek, mint a hormonok (oxitocin, vazopresszin), antibiotikumok stb. A húsznál több aminosavból álló láncot ún. polipeptid, a több mint 60 aminosavból álló polipeptidek pedig fehérjék.

A fehérje szerkezeti szerveződésének szintjei. A fehérjék elsődleges, másodlagos, harmadlagos és kvaterner szerkezetűek lehetnek.
A fehérje elsődleges szerkezete- Ezt lineáris aminosav szekvencia peptidkötés köti össze. Az elsődleges szerkezet végső soron meghatározza a fehérje specificitását és egyediségét, mert még ha feltételezzük is, hogy egy átlagos fehérje 500 aminosavból áll, akkor a lehetséges kombinációk száma 20 500. Ezért legalább egy aminosav helyének változása A sav az elsődleges szerkezetben a másodlagos és magasabb struktúrák, valamint a fehérje egészének tulajdonságainak változását vonja maga után.

A fehérje szerkezeti sajátosságai határozzák meg a térbeli pakolódását - a másodlagos és harmadlagos struktúrák megjelenését.
másodlagos szerkezet a formában lévő fehérjemolekula térbeli elrendezése spirálok vagy redők a hélix vagy redők különböző fordulataiban lévő peptidcsoportok oxigén- és hidrogénatomjai közötti hidrogénkötések tartják. Sok fehérje többé-kevésbé hosszú, másodlagos szerkezetű régiókat tartalmaz. Ilyenek például a haj és köröm keratinjai, a selyemfibroin.
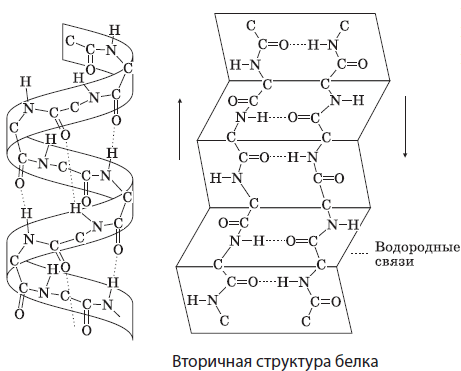
Harmadlagos szerkezet mókus ( gömböcske) szintén a polipeptidlánc térbeli hajtogatásának egy formája, amelyet hidrofób, hidrogén, diszulfid (S-S) és más kötések tartanak fenn. Jellemző a legtöbb testfehérjére, például az izom-mioglobinra.

Negyedidős szerkezet- a legösszetettebb, több polipeptid láncból áll, amelyeket főként ugyanazok a kötések kötnek össze, mint a tercierben (hidrofób, ionos és hidrogén), valamint egyéb gyenge kölcsönhatások. A kvaterner szerkezet néhány fehérjére jellemző, mint például a hemoglobin, a klorofill stb.

A molekula alakja az rostosés gömb alakú fehérjék. Az elsők megnyúltak, mint például a kötőszöveti kollagén vagy a haj- és körömkeratin. A globuláris fehérjék golyó (gömbök) formájában vannak, mint az izom-mioglobin.
Egyszerű és összetett fehérjék. A fehérjék lehetnek egyszerűés összetett. Az egyszerű fehérjék csak aminosavakból állnak, míg összetett fehérjék (lipoproteinek, kromoproteinek, glikoproteinek, nukleoproteinek stb.) tartalmaznak fehérje és nem fehérje részeket. Kromoproteinek színes, nem fehérjetartalmú részt tartalmaznak. Ezek közé tartozik a hemoglobin, a mioglobin, a klorofill, a citokrómok stb. Így a hemoglobin összetételében a globin fehérje mind a négy polipeptid lánca egy nem fehérje részhez kapcsolódik - a hemhez, amelynek középpontjában egy vas található. ion, amely vörös színt ad a hemoglobinnak. Nem fehérje rész lipoproteinek egy lipid és glikoproteinek- szénhidrát. Mind a lipoproteinek, mind a glikoproteinek a sejtmembránok részét képezik. Nukleoproteinek fehérjék és nukleinsavak (DNS és RNS) komplexei. Ők látják el a legfontosabb funkciókat az örökletes információk tárolásának és továbbításának folyamataiban.
A fehérje tulajdonságai. Sok fehérje jól oldódik vízben, de vannak köztük olyanok is, amelyek csak sók, lúgok, savak vagy szerves oldószerek oldatában oldódnak. A fehérjemolekula szerkezete és funkcionális aktivitása a környezeti feltételektől függ. Egy fehérje molekula szerkezetének elvesztését az elsődleges megtartása mellett ún denaturáció.

A denaturáció a hőmérséklet, a pH, a légköri nyomás változása miatt következik be, savak, lúgok, sók hatására nehéz fémek, szerves oldószerek stb. A másodlagos és magasabb struktúrák helyreállításának fordított folyamatát ún renaturáció azonban nem mindig lehetséges. A fehérjemolekula teljes lebomlását ún megsemmisítés.
A fehérje funkciók. A fehérjék számos funkciót látnak el a sejtben: képlékeny (konstrukció), katalitikus (enzimatikus), energia, jel (receptor), összehúzó (motoros), szállító, védő, szabályozó és tárolási funkciót.
A fehérjék építő funkciója a sejtmembránokban és a sejt szerkezeti összetevőiben való jelenlétükkel függ össze. Energia - annak a ténynek köszönhető, hogy 1 g fehérje lebontása során 17,2 kJ energia szabadul fel. A membránreceptor fehérjék aktívan részt vesznek a környezeti jelek érzékelésében és sejten keresztüli továbbításában, valamint az intercelluláris felismerésben. Fehérjék nélkül a sejtek és az organizmusok egészének mozgása lehetetlen, mivel ezek képezik a flagellák és csillók alapját, valamint biztosítják az izomösszehúzódást és az intracelluláris komponensek mozgását. Az emberek és számos állat vérében a hemoglobin fehérje oxigént és szén-dioxid egy részét szállítja, míg más fehérjék ionokat és elektronokat szállítanak. A fehérjék védő szerepe elsősorban az immunitáshoz kapcsolódik, mivel az interferon fehérje számos vírust képes elpusztítani, az antitestfehérjék pedig gátolják a baktériumok és más idegen szerek fejlődését. A fehérjék és peptidek között sok hormon található, például a hasnyálmirigy-hormon, az inzulin, amely szabályozza a vér glükózkoncentrációját. Egyes szervezetekben a fehérjék tartalékban tárolhatók, például a hüvelyesekben a magvakban, vagy a csirke tojás fehérjéiben.
Nukleinsavak
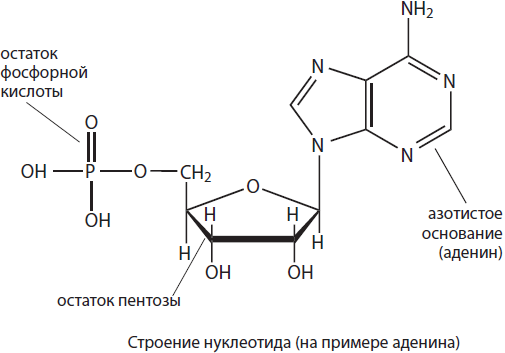
Nukleinsavak olyan biopolimerek, amelyek monomerjei nukleotidok. Jelenleg kétféle nukleinsav ismert: ribonukleinsav (RNS) és dezoxiribonukleinsav (DNS).
Nukleotid egy nitrogéntartalmú bázis, egy pentózcukor- és egy foszforsav-maradék alkotja. A nukleotidok jellemzőit elsősorban az összetételüket alkotó nitrogénbázisok határozzák meg, ezért a nukleotidokat feltételesen is nevük kezdőbetűivel jelöljük. A nukleotidok összetétele öt nitrogénbázist tartalmazhat: adenin (A), guanin (G), timin (T), uracil (U) és citozin (C). A nukleotidok pentózai - ribóz és dezoxiribóz - határozzák meg, hogy melyik nukleotid képződik - ribonukleotid vagy dezoxiribonukleotid. A ribonukleotidok RNS monomerek, jelmolekulákként (cAMP) működhetnek, és részei lehetnek nagy energiájú vegyületeknek, például ATP-nek, valamint koenzimek, mint például NADP, NAD, FAD stb., a dezoxiribonukleotidok pedig a DNS részei.
Dezoxiribonukleinsav (DNS)- kétszálú biopolimer, melynek monomerjei dezoxiribonukleotidok. A dezoxiribonukleotidok összetétele az öt lehetséges közül csak négy nitrogénbázist tartalmaz - adenint (A), timint (T), guanint (G) vagy citozint (C), valamint dezoxiribóz- és foszforsavmaradékot. A DNS-lánc nukleotidjai ortofoszforsav-maradékokon keresztül kapcsolódnak egymáshoz, foszfodiészter kötést képezve. Amikor egy kétszálú molekula képződik, a nitrogéntartalmú bázisok a molekula befelé irányulnak. A DNS-láncok kapcsolódása azonban nem véletlenszerűen történik - a különböző láncok nitrogénbázisait hidrogénkötések kapcsolják össze a komplementaritás elve szerint: az adenint két hidrogénkötés (A \u003d T), valamint guanin és citozin köti össze a timinnel. hárommal (G $ ≡ $ C).

Neki voltak beállítva Feltöltési szabályok:
- Az adenint tartalmazó DNS-nukleotidok száma megegyezik a timint tartalmazó nukleotidok számával (A=T).
- A guanint tartalmazó DNS-nukleotidok száma megegyezik a citozint tartalmazó nukleotidok számával (G$≡$C).
- Az adenint és guanint tartalmazó dezoxiribonukleotidok összege megegyezik a timint és citozint tartalmazó dezoxiribonukleotidok összegével (A+G = T+C).
- Az adenint és timint tartalmazó dezoxiribonukleotidok összegének aránya a guanint és citozint tartalmazó dezoxiribonukleotidok összegéhez viszonyítva a szervezet típusától függ.
A DNS szerkezetét F. Crick és D. Watson fejtette meg ( Nóbel díj in Physiology or Medicine, 1962). Modelljük szerint a DNS-molekula egy jobbkezes kettős hélix. A DNS-lánc nukleotidjai közötti távolság 0,34 nm.

A DNS legfontosabb tulajdonsága a replikációs képesség (önkettőzés). A DNS fő funkciója az örökletes információk tárolása és továbbítása, amelyek nukleotid szekvenciák formájában vannak megírva. A DNS-molekula stabilitását erőteljes javító (helyreállító) rendszerek tartják fenn, de még ezek sem képesek teljesen kiküszöbölni a káros hatásokat, amelyek végső soron mutációkhoz vezetnek. Az eukarióta sejtek DNS-e a sejtmagban, a mitokondriumokban és a plasztidokban koncentrálódik, míg a prokarióta sejtek közvetlenül a citoplazmában találhatók. A nukleáris DNS a kromoszómák alapja, nyílt molekulák képviselik. A mitokondriumok, plasztidok és prokarióták DNS-e kör alakú.
Ribonukleinsav (RNS)- biopolimer, amelynek monomerjei ribonukleotidok. Négy nitrogénbázist is tartalmaznak - adenint (A), uracilt (U), guanint (G) vagy citozint (C), ezáltal eltérnek a DNS-től az egyik bázisban (timin helyett az RNS uracilt tartalmaz). A ribonukleotidokban lévő pentózcukor-maradékot ribóz képviseli. Az RNS többnyire egyszálú molekulák, néhány vírusos molekula kivételével. Az RNS-nek három fő típusa van: információs vagy templát (mRNS, mRNS), riboszómális (rRNS) és transzport (tRNS). Ezek mindegyike a folyamat során keletkezik átiratok- átírás DNS molekulákból.
és Az RNS-ek alkotják a legkisebb RNS-frakciót egy sejtben (2-4%), amit ellensúlyoz a sokféleségük, mivel egy sejt több ezer különböző mRNS-t tartalmazhat. Ezek egyszálú molekulák, amelyek polipeptidláncok szintézisének templátai. A fehérje szerkezetére vonatkozó információkat nukleotidszekvenciák formájában rögzítik bennük, és minden aminosav egy nukleotidhármast kódol - kodon.
R Az RNS a legtöbb RNS típus a sejtben (akár 80%). Molekulatömegük átlagosan 3000-5000; a sejtmagokban képződnek, és a sejtszervecskék - riboszómák - részét képezik. Úgy tűnik, hogy az rRNS-ek szerepet játszanak a fehérjeszintézisben is.
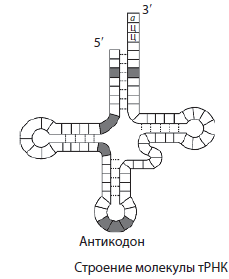
t Az RNS az RNS molekulák közül a legkisebb, mivel mindössze 73-85 nukleotidot tartalmaz. Részesedésük a teljes sejt-RNS-ben körülbelül 16%. A tRNS funkciója az aminosavak szállítása a fehérjeszintézis helyére (riboszómákon). A tRNS-molekula alakja egy lóherelevélre emlékeztet. A molekula egyik végén van egy aminosav kapcsolódási hely, az egyik hurokban pedig egy nukleotidhármas, amely komplementer az mRNS kodonnal, és meghatározza, hogy a tRNS melyik aminosavat hordozza. antikodon.
Az RNS minden típusa aktívan részt vesz az örökletes információ megvalósításában, amely a DNS-ből mRNS-re íródik át, és ez utóbbin történik a fehérjeszintézis. A fehérjeszintézis folyamatában a tRNS aminosavakat szállít a riboszómákhoz, és az rRNS közvetlenül a riboszómák része.
Adenozin-trifoszforsav (ATP) egy nukleotid, amely az adenin nitrogénbázisán és egy ribózcsoporton kívül három foszforsav-maradékot tartalmaz. Az utolsó két foszformaradék közötti kötések makroergikusak (42 kJ/mol energia szabadul fel a hasítás során), míg a szabványos kémiai kötés hasításkor 12 kJ/mol. Ha energiára van szükség, az ATP makroerg kötése felhasad, adenozin-difoszforsav (ADP), foszformaradék képződik, és energia szabadul fel:
ATP + H 2 O $→$ ADP + H 3 PO 4 + 42 kJ.
Az ADP is lebontható AMP-vé (adenozin-monofoszforsav) és foszforsav-maradékká:
ADP + H 2 O $→$ AMP + H 3 PO 4 + 42 kJ.
Az energia-anyagcsere folyamatában (légzés, fermentáció során), valamint a fotoszintézis folyamatában az ADP foszformaradékot köt, és ATP-vé alakul. Az ATP helyreállítási reakcióját nevezzük foszforiláció. Az ATP univerzális energiaforrás az élő szervezetek minden életfolyamatához.
Az összes élő szervezet sejtjeinek kémiai összetételének vizsgálata kimutatta, hogy ugyanazokat a kémiai elemeket tartalmazzák, olyan vegyi anyagokat, amelyek ugyanazokat a funkciókat látják el. Sőt, az egyik szervezetből a másikba átvitt DNS-darab is működni fog benne, a baktériumok vagy gombák által szintetizált fehérje pedig hormonként vagy enzimként működik majd az emberi szervezetben. Ez az egyik bizonyítéka a szerves világ eredetének egységének.

Sejtszerkezet. A sejt épségének alapja a sejtrészek, sejtszervecskék felépítésének és funkcióinak kapcsolata
Sejtszerkezet
A prokarióta és eukarióta sejtek szerkezete
A sejtek fő szerkezeti alkotóelemei a plazmamembrán, a citoplazma és az örökletes apparátus. A szervezet jellemzőitől függően a sejtek két fő típusát különböztetjük meg: prokarióta és eukarióta sejteket. A fő különbség a prokarióta és eukarióta sejtek között az örökletes apparátus felépítése: a prokariótákban közvetlenül a citoplazmában található (a citoplazma ezen területét ún. nukleoid) és nem választják el tőle membránszerkezetek, míg az eukariótákban a DNS nagy része a sejtmagban koncentrálódik, amelyet kettős membrán vesz körül. Emellett a nukleoidban elhelyezkedő prokarióta sejtek genetikai információja a cirkuláris DNS-molekulában rögzítődik, míg az eukariótákban a DNS-molekulák nincsenek zárva.
Az eukariótáktól eltérően a prokarióta sejtek citoplazmája is tartalmaz kis mennyiségben organellumokat, míg az eukarióta sejteket ezeknek a szerkezeteknek a sokfélesége jellemzi.
A biológiai membránok szerkezete és funkciói
A biomembrán szerkezete. Az eukarióta sejtek sejtkötő membránjai és membránszervecskéi közös kémiai összetétellel és szerkezettel rendelkeznek. Ezek közé tartoznak a lipidek, fehérjék és szénhidrátok. A membrán lipidjeit főként foszfolipidek és koleszterin képviselik. A legtöbb membránfehérje összetett fehérje, például glikoproteinek. A szénhidrátok nem fordulnak elő maguktól a membránban, fehérjékhez és lipidekhez kapcsolódnak. A membránok vastagsága 7-10 nm.

A membránszerkezet jelenleg elfogadott folyadékmozaikos modellje szerint a lipidek kettős réteget alkotnak, ill lipid kettős réteg, amelyben a lipidmolekulák hidrofil "fejei" kifelé fordulnak, a hidrofób "farok" pedig a membrán belsejében rejtőzik. Ezek a „farok” hidrofób jellegüknél fogva biztosítják a sejt belső környezetének és környezetének vizes fázisainak elválasztását. A fehérjék különböző típusú kölcsönhatásokon keresztül kapcsolódnak a lipidekhez. A fehérjék egy része a membrán felszínén található. Az ilyen fehérjéket ún kerületi, vagy felszínes. Más fehérjék részben vagy teljesen elmerülnek a membránban – ezek integrál, vagy elmerült fehérjék. A membránfehérjék szerkezeti, transzport, katalitikus, receptor és egyéb funkciókat látnak el.
A membránok nem olyanok, mint a kristályok, alkotóelemeik folyamatosan mozgásban vannak, aminek következtében rések jelennek meg a lipidmolekulák között - pórusok, amelyeken keresztül különféle anyagok juthatnak be vagy távozhatnak a sejtből.
A biológiai membránok különböznek egymástól a sejtben elfoglalt helyükben, kémiai összetételükben és funkcióikban. A membránok fő típusai a plazma és a belső. plazma membrán körülbelül 45% lipidet (beleértve a glikolipidet is), 50% fehérjét és 5% szénhidrátot tartalmaz. A komplex fehérjéket-glikoproteineket és komplex lipideket-glikolipideket alkotó szénhidrátláncok kinyúlnak a membrán felszíne fölé. A plazmaemlős glikoproteinek rendkívül specifikusak. Így például rajtuk keresztül megtörténik a sejtek kölcsönös elismerése, beleértve a spermát és a petéket is.
Az állati sejtek felszínén a szénhidrátláncok vékony felületi réteget alkotnak - glikokalix. Szinte minden állati sejtben megtalálható, de súlyossága nem egyforma (10-50 mikron). A glikokalix közvetlen kapcsolatot biztosít a sejtnek a külső környezettel, extracelluláris emésztés megy végbe benne; receptorok a glikokalixben helyezkednek el. A baktériumok, növények és gombák sejtjeit a plazmalemmán kívül sejtmembránok is körülveszik.
Belső membránok Az eukarióta sejtek elhatárolják a sejt különböző részeit, egyfajta "rekeszeket" alkotva - rekeszek, amely hozzájárul a különböző anyagcsere- és energiafolyamatok szétválasztásához. Kémiai összetételükben és funkciójukban eltérhetnek, de megtartják a szerkezet általános tervét.
A membrán funkciói:
- Korlátozó. Abból áll, hogy elválasztják a sejt belső terét a külső környezettől. A membrán félig áteresztő, vagyis csak azok az anyagok tudnak szabadon legyőzni, amelyek a sejt számára szükségesek, miközben a szükséges anyagok szállítására vannak mechanizmusok.
- Receptor. Elsősorban a környezeti jelek érzékelésével és ezen információknak a sejtbe történő átvitelével kapcsolatos. Különleges receptorfehérjék felelősek ezért a funkcióért. A membránfehérjék a „barát vagy ellenség” elv szerint felelősek a sejtfelismerésért, valamint a sejtközi kapcsolatok kialakításáért is, melyek közül a legtöbbet vizsgált szinapszis. idegsejtek.
- katalitikus. A membránokon számos enzimkomplex található, melynek következtében intenzív szintetikus folyamatok mennek végbe rajtuk.
- Energia átalakító. Az energia képződésével, ATP formájában való tárolásával és kiadásával kapcsolatos.
- Feldarabolás. A membránok a sejten belüli teret is lehatárolják, ezáltal elválasztják a reakció kiindulási anyagait és a megfelelő reakciókat végrehajtani képes enzimeket.
- A sejtek közötti kapcsolatok kialakulása. Annak ellenére, hogy a membrán vastagsága olyan kicsi, hogy szabad szemmel nem lehet megkülönböztetni, egyrészt meglehetősen megbízható gátként szolgál az ionok és molekulák, különösen a vízben oldódó molekulák számára, másrészt , biztosítja azok bejutását a sejtbe és ki.
- Szállítás.

membrán transzport. Annak a ténynek köszönhetően, hogy a sejtek, mint elemi biológiai rendszerek nyílt rendszerek, az anyagcsere és az energia biztosításához, a homeosztázis fenntartásához, a növekedéshez, az ingerlékenységhez és egyéb folyamatokhoz szükséges az anyagok membránon keresztüli átvitele - membrán transzport. Jelenleg az anyagok sejtmembránon keresztül történő szállítása aktív, passzív, endo- és exocitózisra oszlik.
Passzív szállítás egy olyan szállítási mód, amely energiafelhasználás nélkül megy végbe egy magasabb koncentrációból egy alacsonyabb szintre. A lipidben oldódó kis nempoláris molekulák (O 2, CO 2) könnyen behatolnak a sejtbe egyszerű diffúzió. A lipidekben oldhatatlan, beleértve a töltött kis részecskéket is, felveszik a hordozófehérjék, vagy speciális csatornákon (glükóz, aminosavak, K +, PO 4 3-) jutnak át. A passzív szállításnak ezt a fajtáját ún megkönnyített diffúzió. A víz a lipidfázisban lévő pórusokon, valamint speciális fehérjékkel bélelt csatornákon keresztül jut be a sejtbe. A víz membránon keresztül történő szállítását ún ozmózis.

Az ozmózis rendkívül fontos a sejt életében, mert ha nagyobb sókoncentrációjú oldatba helyezzük, mint a sejtoldatban, akkor a víz elkezd elhagyni a sejtet, és az élő tartalom térfogata csökkenni kezd. . Az állati sejtekben a sejt egésze összezsugorodik, a növényi sejtekben pedig a citoplazma lemarad a sejtfal mögött, ami ún. plazmolízis. Ha egy sejtet a citoplazmánál kevésbé koncentrált oldatba helyeznek, a víz az ellenkező irányba - a sejtbe - szállítódik. A citoplazma membrán nyújthatóságának azonban vannak határai, és az állati sejt végül megreped, míg a növényi sejtben ezt az erős sejtfal nem teszi lehetővé. Azt a jelenséget, amikor a sejt teljes belső terét sejttartalommal töltik meg, ún deplazmolízis. Az intracelluláris sókoncentrációt figyelembe kell venni a gyógyszerek elkészítésekor, különösen intravénás adagolás esetén, mivel ez a vérsejtek károsodásához vezethet (ehhez 0,9% nátrium-klorid koncentrációjú sóoldatot használnak). Ez nem kevésbé fontos a sejtek és szövetek, valamint az állatok és növények szerveinek tenyésztésében.

aktiv szállitás az ATP energia felhasználásával egy alacsonyabb koncentrációjú anyagról magasabbra költ. Speciális fehérjepumpák segítségével hajtják végre. A fehérjék K +, Na +, Ca 2+ és más ionokat pumpálnak a membránon keresztül, ami hozzájárul a legfontosabb szerves anyagok szállításához, valamint idegimpulzusok megjelenéséhez stb.
Endocitózis- ez az anyagok sejt általi aktív felszívódásának folyamata, amelyben a membrán invaginációkat, majd membrán hólyagokat képez - fagoszómák, amelyek felszívódott tárgyakat tartalmaznak. Az elsődleges lizoszóma ezután összeolvad a fagoszómával, és kialakul másodlagos lizoszóma, vagy fagolizoszóma, vagy emésztési vakuólum. A vezikula tartalmát a lizoszóma enzimek hasítják, a hasítási termékeket a sejt felszívja és asszimilálja. Az emésztetlen maradványokat exocitózissal távolítják el a sejtből. Az endocitózisnak két fő típusa van: fagocitózis és pinocitózis.
Fagocitózis a sejtfelszín általi befogás és a szilárd részecskék sejt általi felszívódásának folyamata, és pinocytosis- folyadékok. A fagocitózis elsősorban állati sejtekben (egysejtű állatok, emberi leukociták) fordul elő, táplálékukat, gyakran a szervezet védelmét biztosítja. A pinocitózison keresztül megtörténik a fehérjék, az immunreakciók folyamatában antigén-antitest komplexek felszívódása stb.. Sok vírus azonban pinocitózissal vagy fagocitózissal is bejut a sejtbe. A növények és gombák sejtjeiben a fagocitózis gyakorlatilag lehetetlen, mivel erős sejtmembránok veszik körül.

Exocitózis az endocitózis fordított folyamata. Így tűnjön ki emésztetlen maradványok az emésztőüregekből táplálékot, a sejt és a szervezet egészének életéhez szükséges anyagokat eltávolítják. Például az idegimpulzusok átvitele annak köszönhető, hogy az impulzust küldő neuron kémiai hírvivőket bocsát ki - közvetítők, a növényi sejtekben pedig a sejtmembrán segédszénhidrátjai szabadulnak fel ily módon.
Növényi sejtek, gombák és baktériumok sejtfalai. A membránon kívül a sejt erős vázat tud kiválasztani - sejt membrán, vagy sejtfal.
A növényekben a sejtfalat a cellulóz 50-100 molekulát tartalmazó kötegekbe csomagolva. A köztük lévő rések vízzel és más szénhidrátokkal vannak feltöltve. A növényi sejtmembránt tubulusok szúrják át - plazmodezma amelyen áthaladnak az endoplazmatikus retikulum membránjai. A plazmodezmák szállítják az anyagokat a sejtek között. Az anyagok, például a víz szállítása azonban maguk a sejtfalak mentén is megtörténhet. A növények sejtmembránjában idővel különféle anyagok, köztük tanninok vagy zsírszerű anyagok halmozódnak fel, ami magának a sejtfalnak a lignifikációjához vagy eldugulásához, a víz kiszorulásához és a sejttartalom pusztulásához vezet. A szomszédos növényi sejtek sejtfalai között zselészerű párnák vannak - középső lemezek, amelyek rögzítik őket, és cementálják a növény testét egészében. Csak a gyümölcsérés során és a levelek leesésekor pusztulnak el.

Kialakulnak a gombasejtek sejtfalai kitin- nitrogént tartalmazó szénhidrát. Elég erősek és a sejt külső vázát képezik, de a növényekhez hasonlóan megakadályozzák a fagocitózist.
A baktériumokban a sejtfal szénhidrátot tartalmaz peptid-fragmensekkel - murein, azonban tartalma jelentősen eltér a különböző baktériumcsoportokban. A sejtfal tetején más poliszacharidok is felszabadulhatnak, amelyek egy nyálkahártya-kapszulát képeznek, amely megvédi a baktériumokat a külső hatásoktól.
A héj meghatározza a sejt alakját, mechanikai támaszként szolgál, védő funkciót lát el, biztosítja a sejt ozmotikus tulajdonságait, korlátozza az élő tartalom nyúlását és megakadályozza a sejt beáramlása miatt fokozódó felszakadást. víz. Ráadásul a víz és a benne oldott anyagok a citoplazmába jutás előtt, vagy éppen ellenkezőleg, amikor elhagyják a sejtfalat, legyőzik a sejtfalat, miközben a víz gyorsabban szállítódik a sejtfalak mentén, mint a citoplazmán keresztül.
Citoplazma
Citoplazma a sejt belseje. A sejt minden organellumja, a sejtmag és a különféle salakanyagok belemerülnek.
A citoplazma a sejt minden részét összeköti egymással, számos anyagcsere-reakció játszódik le benne. A citoplazmát membránok választják el a környezettől és részekre osztják, vagyis a sejtek membránszerkezettel rendelkeznek. Két halmazállapotú lehet - szol és gél. Sol- ez a citoplazma félig folyékony, zselészerű állapota, amelyben a létfontosságú folyamatok a legintenzívebben mennek végbe, és gél- sűrűbb, kocsonyás állapot, amely akadályozza a kémiai reakciók lefolyását és az anyagok szállítását.
A citoplazma organellumok nélküli folyékony részét ún hialoplazma. A hialoplazma vagy citoszol egy kolloid oldat, amelyben meglehetősen nagy részecskék, például fehérjék egyfajta szuszpenziója van, amelyeket vízmolekulák dipólusai vesznek körül. Ennek a szuszpenziónak az ülepedése nem következik be, mivel azonos töltésűek és taszítják egymást.
Sejtszervecskék
Sejtszervecskék- Ezek a sejt állandó összetevői, amelyek bizonyos funkciókat látnak el.
A szerkezeti jellemzőktől függően membránra és nem membránra osztják őket. Membrán az organellumokat pedig egymembránnak (endoplazmatikus retikulum, Golgi-komplexum és lizoszómák) vagy kettős membránnak (mitokondriumok, plasztidok és mag) nevezik. Nem membrán Az organellumok riboszómák, mikrotubulusok, mikrofilamentumok és a sejtközpont. A felsorolt organellumok közül csak a riboszómák rejlenek a prokariótákban.
A mag felépítése és funkciói. Mag- egy nagy, két membránból álló organellum, amely a sejt közepén vagy a perifériáján fekszik. A mag mérete 3-35 mikron között változhat. A mag alakja gyakrabban gömb vagy ellipszoid, de vannak rúd alakú, orsó alakú, bab alakú, karéjos, sőt tagolt magok is. Egyes kutatók úgy vélik, hogy a sejtmag alakja megfelel magának a sejtnek.
A legtöbb sejtnek egy magja van, de például a máj- és szívsejtekben kettő, számos neuronban pedig akár 15 is lehet. A vázizomrostok általában sok sejtmagot tartalmaznak, de ezek nem a teljes értelemben vett sejtek. a szó, mivel több sejt összeolvadásának eredményeként jönnek létre.
A mag körül van véve sejtmag, és a belső tere megtelt atomlé, vagy nukleoplazma (karioplazma) amelyekbe belemerülnek kromatinés nucleolus. A sejtmag olyan fontos funkciókat lát el, mint az örökletes információk tárolása és továbbítása, valamint a sejtek élettevékenységének szabályozása.

Az acetabularia zöldalgával végzett kísérletek meggyőzően igazolták a mag szerepét az örökletes információk továbbításában. Egyetlen óriási sejtben, amely eléri az 5 cm hosszúságot, kalapot, lábat és rizoidot különböztetnek meg. Ezenkívül csak egy magot tartalmaz, amely a rizoidban található. Az 1930-as években I. Hemmerling az egyik zöld színű acetabularia faj magját egy másik, barna színű rizoidba ültette át, amelyből a magot eltávolították. Egy idő után az átültetett maggal rendelkező növény új sapkát növesztett, mint a mag algadonorája. Ugyanakkor a magot nem tartalmazó rizoidról levált kalap vagy szár egy idő után elpusztult.

sejtmag Két membrán alkotja - külső és belső, amelyek között van egy tér. A membránközi tér a durva endoplazmatikus retikulum üregével kommunikál, a sejtmag külső membránja pedig riboszómákat hordozhat. A nukleáris burkot számos pórus hatja át, és speciális fehérjék szegélyezik. Az anyagok a pórusokon keresztül szállítódnak: a szükséges fehérjék (beleértve az enzimeket is), ionok, nukleotidok és egyéb anyagok bejutnak a sejtmagba, az RNS-molekulák, a hulladékfehérjék, a riboszómák alegységei pedig elhagyják azt. A magburok funkciói tehát a sejtmag tartalmának elválasztása a citoplazmától, valamint a sejtmag és a citoplazma közötti anyagcsere szabályozása.
Nukleoplazma a sejtmag tartalmának nevezzük, amelyben a kromatin és a nukleolusz elmerül. Kolloid oldat, kémiailag a citoplazmára emlékeztet. A nukleoplazma enzimei katalizálják az aminosavak, nukleotidok, fehérjék stb. cseréjét. A nukleoplazma magpórusokon keresztül kapcsolódik a hialoplazmához. A nukleoplazmának a hialoplazmához hasonlóan az a feladata, hogy biztosítsa a sejtmag összes szerkezeti komponensének összekapcsolását és számos enzimatikus reakció végrehajtását.
kromatin vékony szálak és szemcsék halmazának nevezik, amelyek a nukleoplazmába merülnek. Csak festéssel lehet kimutatni, mivel a kromatin és a nukleoplazma törésmutatója megközelítőleg megegyezik. A kromatin fonalas komponensét ún euchromatin, és szemcsés heterokromatin. Az euchromatin gyengén tömörített, mivel örökletes információt olvasnak ki belőle, míg a spiralizált heterokromatin genetikailag inaktív.
A kromatin a kromoszómák szerkezeti módosítása egy nem osztódó magban. Így a kromoszómák folyamatosan jelen vannak a sejtmagban, csak az állapotuk változik attól függően, hogy a mag milyen funkciót lát el.
A kromatin főként nukleoproteinekből (dezoxiribonukleoproteinek és ribonukleoproteinek), valamint enzimekből áll, amelyek közül a legfontosabbak a nukleinsavak szintéziséhez kapcsolódnak, és néhány más anyag.
A kromatin funkciói egyrészt az adott szervezetre specifikus nukleinsavak szintézisében állnak, amelyek specifikus fehérjék szintézisét irányítják, másrészt az örökletes tulajdonságok átvitelében az anyasejtről a leánysejtekre, amelyekhez kromatinszálakat használnak. az osztódás során kromoszómákba csomagolódnak.
nucleolus- gömb alakú test, mikroszkóp alatt jól látható, 1-3 mikron átmérőjű. Az rRNS és a riboszómafehérjék szerkezetére vonatkozó információkat kódoló kromatin régiókban képződik. A sejtmagban a mag gyakran egy, de azokban a sejtekben, ahol intenzív életfolyamatok zajlanak, két vagy több sejtmag is lehet. A sejtmagok funkciója az rRNS szintézise és a riboszóma alegységek összeállítása az rRNS és a citoplazmából származó fehérjék kombinálásával.
Mitokondriumok- kerek, ovális vagy rúd alakú kétmembrán organellumok, bár spirál alakúak is megtalálhatók (a spermiumokban). A mitokondriumok legfeljebb 1 µm átmérőjűek és legfeljebb 7 µm hosszúak. A mitokondriumok belsejében lévő tér tele van mátrixszal. Mátrix Ez a mitokondriumok fő anyaga. Egy kör alakú DNS-molekula és riboszómák merülnek el benne. A mitokondriumok külső membránja sima és sok anyag számára áthatolhatatlan. A belső membrán kinövésekkel rendelkezik - cristae, amelyek megnövelik a membránok felületét a kémiai reakciókhoz. A membrán felületén számos fehérjekomplex található, amelyek az úgynevezett légzési láncot alkotják, valamint gomba alakú ATP-szintetáz enzimek. A mitokondriumokban a légzés aerob szakasza zajlik, melynek során szintetizálódik az ATP.

plasztidok- nagy, két membránból álló organellumok, amelyek csak a növényi sejtekre jellemzőek. A plasztiszok belső tere ki van töltve stroma, vagy mátrix. A stromában többé-kevésbé fejlett membránvezikulák rendszere van - tilakoidok, amelyeket halomba gyűjtenek - gabonafélék, valamint saját, körkörös DNS-molekulája és riboszómái. A plasztidoknak négy fő típusa van: kloroplasztok, kromoplasztok, leukoplasztok és proplasztidok.
Kloroplasztok- Ezek 3-10 mikron átmérőjű, mikroszkóp alatt jól látható zöld plasztiszok. Csak a növények zöld részeiben találhatók - levelekben, fiatal szárban, virágokban és gyümölcsökben. A kloroplasztok többnyire oválisak vagy ellipszoid alakúak, de lehetnek csésze alakúak, spirál alakúak, sőt karéjosak is. A kloroplasztiszok száma egy sejtben átlagosan 10-100 darab. Azonban például egyes algákban ez lehet egy, jelentős méretű és összetett alakú - akkor hívják kromatofor. Más esetekben a kloroplasztiszok száma elérheti a több százat, míg méretük kicsi. A kloroplasztiszok színe a fotoszintézis fő pigmentjének köszönhető - klorofill, bár további pigmenteket tartalmaznak - karotinoidok. A karotinoidok csak ősszel válnak észrevehetővé, amikor az öregedő levelekben lévő klorofill elpusztul. A kloroplasztiszok fő funkciója a fotoszintézis. A fotoszintézis fényreakciói a tilakoid membránokon, amelyeken klorofillmolekulák rögzülnek, a sötét reakciók pedig a számos enzimet tartalmazó stromában mennek végbe.
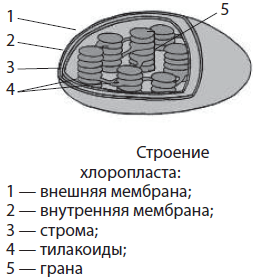
Kromoplasztok sárga, narancssárga és vörös plasztidok, amelyek karotinoid pigmenteket tartalmaznak. A kromoplasztok alakja is jelentősen változhat: cső alakúak, gömb alakúak, kristályosak stb. A kromoplasztok színt adnak a növények virágainak és gyümölcseinek, vonzzák a beporzókat és a magvak és gyümölcsök szétszóródását.
Leukoplasztok- Ezek fehér vagy színtelen plasztiszok, többnyire kerek vagy ovális alakúak. Gyakoriak a nem fotoszintetikus növényrészekben, például levélhéjban, burgonyagumóban stb. Tápanyagokat raktároznak, leggyakrabban keményítőt, de egyes növényekben ez lehet fehérje vagy olaj is.
A plasztidok növényi sejtekben proplasztidokból képződnek, amelyek már jelen vannak a nevelési szövet sejtjeiben, és kis, kétmembrános testek. A korai szakaszaiban fejlődésében a különböző típusú plasztidok képesek egymásba fordulni: fény hatására a burgonyagumó leukoplasztjai és a sárgarépa gyökér kromoplasztjai zöldellnek.
A plasztidokat és a mitokondriumokat félautonóm sejtszervecskéknek nevezzük, mivel saját DNS-molekulájuk és riboszómájuk van, fehérjeszintézist hajtanak végre és a sejtosztódástól függetlenül osztódnak. Ezeket a tulajdonságokat az egysejtű prokarióta szervezetektől való származás magyarázza. A mitokondriumok és a plasztidok "függetlensége" azonban korlátozott, mivel DNS-ük túl kevés gént tartalmaz a szabad létezéshez, míg a többi információ a sejtmag kromoszómáiban van kódolva, ami lehetővé teszi ezen organellumok irányítását.
Endoplazmatikus retikulum (ER), vagy endoplazmatikus retikulum (ER), egy egymembránból álló organellum, amely membránüregek és tubulusok hálózata, amely a citoplazma tartalmának akár 30%-át is elfoglalja. Az ER tubulusok átmérője körülbelül 25-30 nm. Kétféle EPS létezik - durva és sima. Durva XPS riboszómákat hordoz, és itt szintetizálódnak a fehérjék. Sima EPS riboszómáktól mentes. Feladata a lipidek és szénhidrátok szintézise, valamint a mérgező anyagok szállítása, tárolása és ártalmatlanítása. Kifejezetten azokban a sejtekben fejlődik ki, ahol intenzív anyagcsere-folyamatok mennek végbe, például a májsejtekben - hepatocitákban - és a vázizomrostokban. Az EPS-ben szintetizált anyagok a Golgi-készülékbe kerülnek. Az ER-ben a sejtmembránok is összeállnak, de kialakulásuk a Golgi-készülékben fejeződik be.
golgi készülék, vagy golgi komplexum, lapos ciszternák, tubulusok és a belőlük leszakadt hólyagok rendszeréből kialakított egymembrán organellum. A Golgi-apparátus szerkezeti egysége az diktaszóma- egy köteg tartály, amelynek egyik pólusára az ER-ből származó anyagok jönnek, a másik pólusról pedig bizonyos átalakulások után buborékokba csomagolják és a cella más részeibe küldik. A tartályok átmérője körülbelül 2 mikron, a kis buborékok körülbelül 20-30 mikron. A Golgi komplex fő funkciói bizonyos anyagok szintézise és az ER-ből érkező fehérjék, lipidek és szénhidrátok módosítása (megváltoztatása), a membránok végső kialakítása, valamint az anyagok sejten keresztüli szállítása, szerkezetét és a lizoszómák képződését. A Golgi-készülék nevét Camillo Golgi olasz tudós tiszteletére kapta, aki először fedezte fel ezt az organoidot (1898).

Lizoszómák- kisméretű, legfeljebb 1 mikron átmérőjű egymembrán organellumok, amelyek az intracelluláris emésztésben részt vevő hidrolitikus enzimeket tartalmaznak. A lizoszómák membránja gyengén permeábilis ezen enzimek számára, így funkcióik lizoszómák általi ellátása nagyon pontos és célzott. Tehát aktívan részt vesznek a fagocitózis folyamatában, emésztési vakuolák kialakításában, és éhezés vagy a sejt bizonyos részeinek károsodása esetén megemésztik azokat anélkül, hogy másokat befolyásolnának. Az utóbbi időben felfedezték a lizoszómák szerepét a sejthalál folyamataiban.
Vacuole- üreg a növényi és állati sejtek citoplazmájában, amelyet membrán határol és folyadékkal töltenek meg. Emésztőrendszeri és kontraktilis vakuolák találhatók a protozoon sejtekben. Előbbiek részt vesznek a fagocitózis folyamatában, mivel lebontják a tápanyagokat. Ez utóbbiak az ozmoreguláció révén biztosítják a víz-só egyensúly fenntartását. Többsejtű állatokban főként emésztési vakuolák találhatók.
A növényi sejtekben a vakuolák mindig jelen vannak, ezeket speciális membrán veszi körül, és sejtnedvvel töltik meg. A vakuólumot körülvevő membrán kémiai összetételében, szerkezetében és funkciójában hasonló a plazmamembránhoz. sejtnedv Különféle szervetlen és szerves anyagok vizes oldatát képviseli, beleértve az ásványi sókat, szerves savakat, szénhidrátokat, fehérjéket, glikozidokat, alkaloidokat stb. A vakuólum a sejttérfogat 90%-át is elfoglalhatja, és a magot a perifériára szorítja. A sejtnek ez a része raktározó, kiválasztó, ozmotikus, védő, lizoszómális és egyéb funkciókat lát el, hiszen tápanyagokat és salakanyagokat halmoz fel, vízellátást biztosít, fenntartja a sejt alakját és térfogatát, emellett számos sejt lebontásához szükséges enzimeket is tartalmaz. sejtkomponensek. Ráadásul biológiailag hatóanyagok vakuolák képesek megakadályozni, hogy sok állat elfogyassza ezeket a növényeket. Számos növényben a vakuólumok duzzanata miatt a sejtnövekedés nyújtással megy végbe.
A vakuolák egyes gombák és baktériumok sejtjeiben is jelen vannak, de a gombákban csak az ozmoreguláció funkcióját látják el, míg a cianobaktériumokban fenntartják a felhajtóerőt és részt vesznek a levegőből származó nitrogén asszimiláció folyamataiban.
Riboszómák- 15-20 mikron átmérőjű kis nem membrán organellumok, amelyek két alegységből állnak - nagy és kicsi. Az eukarióta riboszóma alegységek a sejtmagban állnak össze, majd a citoplazmába szállítják. A prokarióták, mitokondriumok és plasztidok riboszómái kisebbek, mint az eukariótáké. A riboszóma alegységei közé tartoznak az rRNS és a fehérjék.

Egy sejtben a riboszómák száma elérheti a több tízmilliót is: a citoplazmában, a mitokondriumokban és a plasztidokban szabad állapotban, a durva ER-en pedig kötött állapotban vannak. Részt vesznek a fehérjeszintézisben, különösen a transzlációs folyamatot - egy polipeptidlánc bioszintézisét egy mRNS-molekulán. A szabad riboszómákon a hialoplazma, a mitokondriumok, a plasztidok és a riboszómák saját fehérjéi szintetizálódnak, míg a durva ER-hez kapcsolódó riboszómákon a fehérjék transzlációja történik a sejtekből való kiválasztódás, a membránok összeállítása, a lizoszómák és vakuolák képződése érdekében.
A riboszómák elhelyezkedhetnek a hialoplazmában egyenként vagy csoportokba rendezve, több polipeptid lánc egyidejű szintézisével egy mRNS-en. Ezeket a riboszómacsoportokat ún poliriboszómák, vagy poliszómák.

mikrotubulusok- Ezek hengeres üreges nem membránszervek, amelyek a sejt teljes citoplazmáján áthatolnak. Átmérőjük kb. 25 nm, falvastagságuk 6-8 nm. Számos fehérjemolekulából állnak. tubulin, amelyek először 13 gyöngyökre emlékeztető szálat alkotnak, majd mikrotubulussá állnak össze. A mikrotubulusok citoplazmatikus retikulumot alkotnak, amely a sejt alakját és térfogatát adja, összeköti a plazmamembránt a sejt többi részével, biztosítja az anyagok sejten keresztüli szállítását, részt vesz a sejt és az intracelluláris komponensek mozgásában, valamint az osztódásban. genetikai anyag. Részei a sejtközpontnak és a mozgásszervek - flagellák és csillók.
mikrofilamentumok, vagy mikrofilamentumok, szintén nem membrán organellumok, azonban fonalas alakúak, és nem tubulin, hanem aktinóma. Részt vesznek a membrántranszport folyamataiban, az intercelluláris felismerésben, a sejt citoplazma osztódásában és mozgásában. Az izomsejtekben az aktin mikrofilamentumok és a miozin filamentumok kölcsönhatása biztosítja az összehúzódást.
Mikrotubulusok és mikrofilamentumok alkotják a sejt belső vázát citoszkeleton. Összetett rosthálózat, amely a plazmamembrán mechanikai alátámasztását biztosítja, meghatározza a sejt alakját, a sejtszervecskék elhelyezkedését és mozgásukat a sejtosztódás során.

Cell Center- nem membrán organellum, amely állati sejtekben található a sejtmag közelében; a növényi sejtekben hiányzik. Hossza körülbelül 0,2–0,3 µm, átmérője 0,1–0,15 µm. A sejtközpont kettőből áll centriolák egymásra merőleges síkban fekvő, és sugárzó gömb mikrotubulusokból. Mindegyik centriólum kilenc mikrotubuluscsoportból áll, amelyeket hármasban, azaz hármasban gyűjtenek össze. A sejtközpont részt vesz a mikrotubulusok összeállításában, a sejt örökítőanyagának osztódásában, valamint a flagellák és csillók képzésében.

A mozgás szervei. Flagellaés csillók plazmalemmával borított sejtek kinövései. Ezek az organellumok kilenc pár mikrotubuluson alapulnak, amelyek a periférián helyezkednek el, és két szabad mikrotubuluson a központban. A mikrotubulusokat különféle fehérjék kötik össze, amelyek biztosítják a tengelytől való koordinált eltérést - oszcillációt. A fluktuációk energiafüggőek, vagyis az ATP makroerg kötéseinek energiáját fordítják erre a folyamatra. Az elveszett flagellák és csillók helyreállítása funkció bazális testek, vagy kinetoszómák a bázisukon találhatók.
A csillók hossza körülbelül 10-15 nm, a flagella hossza 20-50 mikron. A flagellák és csillók szigorúan irányított mozgása miatt nemcsak az egysejtű állatok, spermiumok stb. mozgása történik meg, hanem a légutak is kitisztulnak, a pete együtt mozog petevezetékek, hiszen az emberi test mindezen részeit csillós hám borítja.

Zárványok
Zárványok- Ezek a sejt nem állandó alkotórészei, amelyek élete során keletkeznek és eltűnnek. Ide tartoznak a tartalék anyagok, például a keményítő- vagy fehérjeszemek a növényi sejtekben, a glikogén szemcsék állati és gombás sejtekben, a volutin baktériumokban, a zsírcseppek minden sejttípusban, valamint a salakanyagok, különösen az ebből eredő emésztetlen élelmiszer-maradványok. fagocitózis, úgynevezett maradéktesteket képezve.

A sejt épségének alapja a sejtrészek, sejtszervecskék felépítésének és funkcióinak kapcsolata
A sejt egyes részei egyrészt különálló szerkezetek, amelyek sajátos felépítésű és funkciójúak, másrészt egy további elem alkotóeleme. összetett rendszer sejtnek nevezik. Az eukarióta sejt örökletes információinak nagy része a sejtmagban koncentrálódik, de a sejtmag maga nem tudja biztosítani a megvalósítását, hiszen ehhez legalább a fő anyagként működő citoplazmára és riboszómákra van szükség, amelyeken ez a szintézis megtörténik. . A legtöbb riboszóma a szemcsés endoplazmatikus retikulumon helyezkedik el, ahonnan a fehérjék leggyakrabban a Golgi komplexbe kerülnek, majd módosítás után a sejt azon részeire, amelyekre szánják, illetve kiürülnek. A fehérjék és szénhidrátok membrános csomagolása szerves membránokba és citoplazmatikus membránokba integrálható, biztosítva azok folyamatos megújulását. A legfontosabb funkciókat ellátó lizoszómákat és vakuolákat is a Golgi-komplexumból csipkézik. Például lizoszómák nélkül a sejtek gyorsan a hulladékmolekulák és -struktúrák egyfajta lerakóhelyévé válnának.
Mindezekhez a folyamatokhoz a mitokondriumok és növényekben a kloroplasztiszok által termelt energiára van szükség. És bár ezek az organellumok viszonylag önállóak, mivel saját DNS-molekulájuk van, egyes fehérjéket még mindig a nukleáris genom kódol, és a citoplazmában szintetizálódik.
Így a sejt az alkotóelemeinek elválaszthatatlan egysége, amelyek mindegyike ellátja a maga egyedi funkcióját.
Az anyagcsere és az energiaátalakítás az élő szervezetek tulajdonságai. Energia- és képlékeny anyagcsere, kapcsolatuk. Az energia-anyagcsere szakaszai. Erjedés és légzés. Fotoszintézis, jelentősége, kozmikus szerepe. A fotoszintézis fázisai. A fotoszintézis világos és sötét reakciói, kapcsolatuk. Kemoszintézis. A kemoszintetikus baktériumok szerepe a Földön
Anyagcsere és energiaátalakítás - az élő szervezetek tulajdonságai
A sejt egy miniatűr vegyi gyárhoz hasonlítható, ahol kémiai reakciók százai és ezrei játszódnak le.
Anyagcsere- a biológiai rendszerek megőrzését és önszaporodását célzó kémiai átalakulások összessége.
Ide tartozik a táplálkozás és a légzés során a szervezetbe jutó anyagok, az intracelluláris anyagcsere, ill anyagcsere, valamint az anyagcsere végtermékeinek kiosztása.
Az anyagcsere elválaszthatatlanul összefügg az egyik energiafajtának a másikká történő átalakításának folyamataival. Például a fotoszintézis folyamatában a fényenergiát összetett szerves molekulák kémiai kötéseinek energiája formájában tárolják, és a légzés során felszabadul, és új molekulák szintézisére, mechanikai és ozmotikus munkára fordítják, hő formájában eloszlik stb.
Az élő szervezetekben a kémiai reakciók lefolyását fehérje természetű biológiai katalizátorok biztosítják - enzimek, vagy enzimek. Más katalizátorokhoz hasonlóan az enzimek a sejtben a kémiai reakciók lefolyását tíz- és százezerszeresére gyorsítják, sőt olykor lehetővé is teszik, de nem változtatják meg sem a reakció végtermékének (termékeinek) jellegét, sem tulajdonságait, ne változtassák meg magukat. Az enzimek lehetnek egyszerű és összetett fehérjék is, amelyek a fehérje részen kívül tartalmaznak nem fehérje részt is - kofaktor (koenzim). Az enzimekre példa a nyál amiláz, amely a poliszacharidokat bontja le hosszan tartó rágás során, és a pepszin, amely biztosítja a fehérjék emésztését a gyomorban.
Az enzimek abban különböznek a nem fehérje katalizátoroktól, hogy hatásuk magas, a segítségükkel jelentősen megnövekszik a reakciósebesség, valamint képesek szabályozni a hatást a reakciókörülmények megváltoztatásával vagy különféle anyagokkal való kölcsönhatásban. Ezenkívül az enzimatikus katalízis körülményei jelentősen eltérnek a nem enzimatikus katalízistől: a 37 °C USD hőmérséklet optimális az enzimek működéséhez az emberi szervezetben, a nyomásnak közel kell lennie a légköri nyomáshoz, és a közeg $pH$-ja jelentősen tétovázhat. Tehát az amilázhoz lúgos környezet szükséges, a pepszinhez pedig savas.
Az enzimek hatásmechanizmusa a reakcióba belépő anyagok (szubsztrátok) aktiválási energiájának csökkentése a közbenső enzim-szubsztrát komplexek képződése miatt.

Energia- és képlékeny anyagcsere, kapcsolatuk
Az anyagcsere a sejtben egyidejűleg két folyamatból áll: képlékeny és energiacseréből.
Plasztikus anyagcsere (anabolizmus, asszimiláció) szintézisreakciók összessége, amelyek az ATP energia felhasználásával járnak. A képlékeny anyagcsere folyamatában a sejt számára szükséges szerves anyagok szintetizálódnak. A képlékeny cserereakciók példái a fotoszintézis, a fehérjebioszintézis és a DNS-replikáció (önkettőzés).
Energiaanyagcsere (katabolizmus, disszimiláció) olyan reakciók összessége, amelyek az összetett anyagokat egyszerűbbekre bontják. Az energia-anyagcsere eredményeként energia szabadul fel, ATP formájában raktározódik. Az energiaanyagcsere legfontosabb folyamatai a légzés és az erjedés.
A műanyag és az energiacsere elválaszthatatlanul összefügg, mivel a képlékeny csere folyamatában szerves anyagok szintetizálódnak, és ehhez ATP energiára van szükség, az energiaanyagcsere folyamatában pedig a szerves anyagok széthasadnak és energia szabadul fel, amit aztán a szintézis folyamatokra fordítanak. .
Az élőlények a táplálkozás során energiát kapnak, és főként a légzés során felszabadítják és hozzáférhető formává alakítják. A táplálkozás módja szerint minden élőlény autotrófokra és heterotrófokra oszlik. Autotrófok képes önállóan szintetizálni szerves anyagokat szervetlenből, és heterotrófok kizárólag kész szerves anyagokat használjon.
Az energia-anyagcsere szakaszai
Az energia-anyagcsere-reakciók összetettsége ellenére feltételesen három szakaszra oszlik: előkészítő, anaerob (oxigénmentes) és aerob (oxigén) szakaszra.
A előkészítő szakasz a poliszacharidok, lipidek, fehérjék, nukleinsavak egyszerűbb molekuláira bomlanak le, például glükózra, glicerinre és zsírsavakká, aminosavakká, nukleotidokká stb. Ez a szakasz végbemehet közvetlenül a sejtekben vagy a bélben, ahonnan a a hasított anyagokat vérárammal szállítják.
anaerob szakasz Az energiametabolizmust a szerves vegyületek monomereinek további hasadása kíséri még egyszerűbb köztes termékekké, például piroszőlősavvá vagy piruváttá. Nem igényel oxigén jelenlétét, és sok, a mocsarak iszapjában vagy az emberi bélrendszerben élő szervezet számára az egyetlen módja energiát kapni. Az energia-anyagcsere anaerob szakasza a citoplazmában zajlik.
Különféle anyagok oxigénmentesen hasíthatnak, de gyakran a glükóz a reakciók szubsztrátja. Oxigénmentes felhasadásának folyamatát ún glikolízis. A glikolízis során a glükózmolekula négy hidrogénatomot veszít, azaz oxidálódik, és két molekula piroszőlősav, két ATP molekula és két redukált hidrogénhordozó molekula $NADH + H^(+)$ keletkezik:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Az ATP képződése az ADP-ből egy foszfát anionnak egy korábban foszforilált cukorból való közvetlen átvitele miatt következik be, és az ún. szubsztrát foszforiláció.
Aerobik színpad energiacsere csak oxigén jelenlétében mehet végbe, miközben az oxigénmentes hasadás során keletkező köztes vegyületek végtermékekké (szén-dioxid és víz) oxidálódnak, és a szerves vegyületek kémiai kötéseiben tárolt energia nagy része felszabadul. . 36 ATP molekula makroerg kötéseinek energiájába megy át. Ezt a szakaszt más néven szöveti légzés. Oxigén hiányában a köztes vegyületek más szerves anyagokká alakulnak, ezt a folyamatot ún erjesztés.
Lehelet
A sejtlégzés mechanizmusát vázlatosan az 1. ábra mutatja.

Az aerob légzés a mitokondriumokban megy végbe, míg a piroszőlősav először egy szénatomot veszít, ami egy redukáló ekvivalens $NADH + H^(+)$ és egy acetil-koenzim A (acetil-CoA) molekula szintézisével jár együtt:
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
A mitokondriális mátrixban lévő acetil-CoA kémiai reakciók láncolatában vesz részt, amelyek összességét ún. Krebs ciklus (trikarbonsav ciklus, citromsav ciklus). Ezen átalakulások során két ATP molekula keletkezik, az acetil-CoA teljesen szén-dioxiddá oxidálódik, hidrogénionjai és elektronjai a $NADH + H^(+)$ és $FADH_2$ hidrogénhordozókhoz kapcsolódnak. A hordozók hidrogén protonokat és elektronokat szállítanak a mitokondriumok belső membránjaihoz, amelyek krisztákat képeznek. A hordozófehérjék segítségével a hidrogén protonokat injektálják a membránközi térbe, az elektronok pedig az ún. légzőlánc A mitokondriumok belső membránján található enzimek, amelyek az oxigénatomokra kerülnek:
$O_2+2e^(-)→O_2^-$.
Meg kell jegyezni, hogy a légzőlánc egyes fehérjéi vasat és ként tartalmaznak.
A membránközi térből speciális enzimek - ATP-szintázok - segítségével a hidrogén protonok visszajutnak a mitokondriális mátrixba, és az ilyenkor felszabaduló energiát minden glükózmolekulából 34 ATP-molekula szintézisére fordítják. Ezt a folyamatot ún oxidatív foszforiláció. A mitokondriális mátrixban a hidrogén protonok reakcióba lépnek az oxigéngyökökkel, és vizet képeznek:
$4H^(+)+O_2^-→2H_2O$.
Az oxigénlégzés reakcióinak halmaza a következőképpen fejezhető ki:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
A teljes légzési egyenlet így néz ki:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Erjesztés
Oxigén hiányában vagy annak hiányában erjedés következik be. Az erjesztés evolúciósan korábbi módja az energiaszerzésnek, mint a légzés, de energetikailag kevésbé kifizetődő, mivel az erjesztés során még energiadús szerves anyagok keletkeznek. Az erjesztésnek több fő típusa van: tejsav, alkohol, ecetsav stb.. Tehát a vázizmokban az erjedés során oxigén hiányában a piroszőlősav tejsavvá redukálódik, miközben a korábban képződött redukáló ekvivalensek elfogynak, ill. csak két ATP-molekula maradt:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Az élesztőgombák segítségével történő erjesztés során a piroszőlősav oxigén jelenlétében etil-alkohollá és szén-monoxiddá (IV) alakul:
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
A mikroorganizmusok segítségével történő erjesztés során a piroszőlősavból ecetsav, vajsav, hangyasav stb.
Az energia-anyagcsere eredményeként nyert ATP-t a sejtben különféle típusú munkákhoz használjuk fel: kémiai, ozmotikus, elektromos, mechanikai és szabályozó. A kémiai munka fehérjék, lipidek, szénhidrátok, nukleinsavak és más létfontosságú vegyületek bioszintéziséből áll. Az ozmotikus munka magában foglalja a sejt általi felszívódási folyamatokat és az olyan anyagok eltávolítását, amelyek az extracelluláris térben nagyobb koncentrációban vannak, mint magában a sejtben. Az elektromos munka szorosan összefügg az ozmotikus munkával, mivel a töltött részecskék membránokon keresztüli mozgása következtében alakul ki a membrán töltése, sajátítják el a gerjeszthetőség és vezetőképesség tulajdonságait. A mechanikai munka az anyagok és szerkezetek sejten belüli mozgásával, valamint a sejt egészével kapcsolatos. A szabályozási munka magában foglal minden olyan folyamatot, amely a sejtben zajló folyamatok koordinálására irányul.
Fotoszintézis, jelentősége, kozmikus szerepe
fotoszintézis a fényenergiát a szerves vegyületek kémiai kötéseinek energiájává alakító folyamatnak nevezik klorofill részvételével.
A fotoszintézis eredményeként évente mintegy 150 milliárd tonna szerves anyag és hozzávetőleg 200 milliárd tonna oxigén keletkezik. Ez a folyamat biztosítja a szén keringését a bioszférában, megakadályozva a szén-dioxid felhalmozódását, és ezáltal az üvegházhatás kialakulását és a Föld túlmelegedését. A fotoszintézis eredményeként keletkező szerves anyagokat más élőlények nem fogyasztják el teljesen, jelentős részük évmilliók alatt ásványi lerakódásokat (kemény- és barnaszén, olaj) képez. NÁL NÉL mostanában a repceolajat („biodízelt”) és a növényi maradványokból nyert alkoholt is elkezdték üzemanyagként használni. Az oxigénből elektromos kisülések hatására ózon képződik, amely ózonpajzsot képez, amely megvédi a Föld minden élővilágát az ultraibolya sugárzás káros hatásaitól.
Honfitársunk, a kiváló növényfiziológus, K. A. Timirjazev (1843-1920) „kozmikusnak” nevezte a fotoszintézis szerepét, mivel összeköti a Földet a Nappal (űrrel), energia beáramlást biztosítva a bolygó számára.
A fotoszintézis fázisai. A fotoszintézis világos és sötét reakciói, kapcsolatuk
1905-ben F. Blackman angol növényfiziológus felfedezte, hogy a fotoszintézis sebessége nem nőhet a végtelenségig, bizonyos tényezők korlátozzák. Ennek alapján a fotoszintézis két fázisának létezését javasolta: könnyűés sötét. Alacsony fényintenzitás esetén a fényreakciók sebessége a fényintenzitás növekedésével arányosan növekszik, ráadásul ezek a reakciók nem függnek a hőmérséklettől, mivel nem igényelnek enzimeket. Fényreakciók lépnek fel a tilakoid membránokon.
A sötét reakciók sebessége ezzel szemben a hőmérséklet emelkedésével növekszik, azonban a 30°C$-os hőmérsékleti küszöb elérésekor ez a növekedés leáll, ami jelzi a stromában végbemenő átalakulások enzimatikus jellegét. Meg kell jegyezni, hogy a fény bizonyos hatást gyakorol a sötét reakciókra is, annak ellenére, hogy ezeket sötétnek nevezik.

A fotoszintézis könnyű fázisa tilakoid membránokon megy végbe, amelyek többféle fehérjekomplexet hordoznak, amelyek közül a legfontosabb az I. és II. fotorendszer, valamint az ATP-szintáz. A fotorendszerek összetétele pigment komplexeket tartalmaz, amelyekben a klorofill mellett karotinoidok is vannak. A karotinoidok megfogják a fényt a spektrum azon részein, ahol a klorofill nem, és megvédi a klorofillt a nagy intenzitású fény által okozott pusztulástól.
A pigmentkomplexeken kívül a fotorendszerek számos elektronakceptor fehérjét is tartalmaznak, amelyek egymás után adják át az elektronokat a klorofillmolekulákról. Ezeknek a fehérjéknek a sorrendjét ún kloroplaszt elektrontranszport lánc.
A II. fotorendszerhez egy speciális fehérjekomplex is társul, amely biztosítja az oxigén felszabadulását a fotoszintézis során. Ez az oxigénfejlesztő komplex mangán- és klórionokat tartalmaz.
NÁL NÉL világos fázis a tilakoid membránokon elhelyezkedő klorofill molekulákra eső fénykvantumok vagy fotonok gerjesztett állapotba juttatják őket, amelyekre több jellemző nagy energia elektronok. Ugyanakkor az I. fotorendszer klorofilljából gerjesztett elektronok egy közvetítő láncon keresztül a hidrogénhordozó NADP-hez jutnak, amely hidrogén-protonokat ad hozzá, amelyek mindig jelen vannak a vizes oldatban:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
A csökkentett $NADPH + H^(+)$ ezután a sötét szakaszban lesz felhasználva. A II. fotoszisztéma klorofilljából származó elektronok is átkerülnek az elektrontranszport láncon, de ezek kitöltik az I. fotorendszer klorofillában lévő „elektronlyukakat”. molekulák, ami a fent már említett oxigén-leadó komplex részvételével történik. A vízmolekulák bomlásának eredményeként, amelyet ún fotolízis hidrogén protonok keletkeznek és molekuláris oxigén szabadul fel, ami a fotoszintézis mellékterméke:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Genetikai információ a sejtben. Gének, genetikai kód és tulajdonságai. Bioszintetikus reakciók mátrix jellege. Fehérje és nukleinsavak bioszintézise
Genetikai információ a sejtben
A saját fajták szaporodása az élők egyik alapvető tulajdonsága. Ennek a jelenségnek köszönhetően nemcsak az élőlények között van hasonlóság, hanem az egyes sejtek, illetve azok organellumjai (mitokondriumok és plasztidok) között is. Ennek a hasonlóságnak az anyagi alapja a DNS-nukleotid szekvenciában titkosított genetikai információ továbbítása, amely a DNS-replikáció (önkettőzés) folyamatai miatt valósul meg. A sejtek és élőlények minden jellemzője és tulajdonsága a fehérjéknek köszönhetően valósul meg, amelyek szerkezetét elsősorban a DNS nukleotid szekvenciák határozzák meg. Ezért az anyagcsere folyamatokban a nukleinsavak és fehérjék bioszintézise kiemelten fontos. Az öröklődő információ szerkezeti egysége a gén.
Gének, genetikai kód és tulajdonságai
A sejtben lévő örökletes információ nem monolitikus, külön "szavakra" - génekre - van osztva.
Gén a genetikai információ alapegysége.
Az „Emberi genom” programon végzett munka, amelyet több országban egyidejűleg végeztek, és a század elején fejezték be, megértettük, hogy az embernek csak körülbelül 25-30 ezer génje van, de a legtöbbünk információi. A DNS-t soha nem olvassák ki, mivel rengeteg értelmetlen szakaszt, ismétlődést és olyan gént tartalmaz, amelyek olyan tulajdonságokat kódolnak, amelyek az ember számára értelmüket vesztették (farok, testszőrzet stb.). Emellett számos örökletes betegségek kialakulásáért felelős gént, valamint gyógyszer célgéneket sikerült megfejteni. A program megvalósítása során kapott eredmények gyakorlati alkalmazása azonban elodázódik, amíg több ember genomját dekódolják, és kiderül, miben különböznek egymástól.
A fehérje elsődleges szerkezetét, riboszómális vagy transzfer RNS-t kódoló géneket nevezzük szerkezetiés olyan gének, amelyek aktiválják vagy elnyomják a szerkezeti génekből származó információolvasást, szabályozó. Azonban még a szerkezeti gének is tartalmaznak szabályozó régiókat.
Az élőlények örökletes információi a DNS-ben titkosítva vannak bizonyos nukleotid-kombinációk és azok szekvenciája formájában - genetikai kód. Tulajdonságai: triplet, specifitás, univerzalitás, redundancia és átfedésmentesség. Ráadásul a genetikai kódban nincsenek írásjelek.
Minden aminosavat három nukleotid kódol a DNS-ben. hármas például a metionint a TAC triplet, azaz a triplet kód kódolja. Másrészt minden hármas csak egy aminosavat kódol, ami annak specifitása vagy egyértelműsége. A genetikai kód minden élő szervezet számára univerzális, vagyis az emberi fehérjékről származó örökletes információkat a baktériumok kiolvashatják és fordítva. Ez a szerves világ eredetének egységéről tanúskodik. Három nukleotidból álló 64 kombináció azonban csak 20 aminosavnak felel meg, aminek következtében 2-6 triplett kódolhat egy aminosavat, vagyis a genetikai kód redundáns, vagy degenerált. Három hármasnak nincs megfelelő aminosavja, ezeket hívják stop kodonok, mivel ezek jelzik a polipeptid lánc szintézisének végét.
A bázisok sorrendje a DNS-hármasokban és az általuk kódolt aminosavak

*Stop kodon, amely a polipeptid lánc szintézisének végét jelzi.
Az aminosavnevek rövidítései:
Ala - alanin
Arg - arginin
Asn - aszparagin
Asp - aszparaginsav
Val - valin
Az ő - hisztidin
Gly - glicin
Gln – glutamin
Glu - glutaminsav
Ile - izoleucin
Leu - leucin
Liz - lizin
Meth - metionin
Pro - prolin
Ser - szerin
Tyr - tirozin
Tre - treonin
Három - triptofán
Fen - fenilalanin
cisz-cisztein
Ha a genetikai információt nem a hármasban lévő első nukleotidtól kezdi el olvasni, hanem a másodiktól, akkor nemcsak az olvasási keret fog eltolódni - az így szintetizált fehérje nemcsak a nukleotidszekvenciában, hanem szerkezetében is teljesen más lesz. és tulajdonságait. A tripletek között nincsenek írásjelek, így nincs akadálya az olvasási keret eltolódásának, ami teret nyit a mutációk előfordulásának és fenntartásának.
Bioszintetikus reakciók mátrix jellege
A bakteriális sejtek 20-30 percenként képesek megkettőzni, az eukarióta sejtek pedig minden nap és még gyakrabban, amihez a DNS-replikáció nagy sebessége és pontossága szükséges. Ráadásul minden sejt több száz és ezer kópiát tartalmaz számos fehérjéből, különösen enzimekből, ezért szaporodásuk szempontjából elfogadhatatlan a "darabos" előállítási módszerük. Egy progresszívebb módszer a bélyegzés, amely lehetővé teszi, hogy számos pontos másolatot kapjon a termékről, és csökkentse annak költségeit. A bélyegzéshez mátrixra van szükség, amellyel lenyomatot készítenek.
A sejtekben a mátrix szintézis elve az, hogy új fehérjék és nukleinsavmolekulák szintetizálódnak az azonos nukleinsavak (DNS vagy RNS) már meglévő molekuláinak szerkezetében lefektetett program szerint.
Fehérje és nukleinsavak bioszintézise
DNS replikáció. A DNS egy kétszálú biopolimer, amelynek monomerjei nukleotidok. Ha a DNS-bioszintézis a fénymásolás elve szerint zajlana, akkor elkerülhetetlenül számos torzulás és hiba keletkezne az öröklődő információkban, amelyek végső soron új élőlények pusztulásához vezetnének. Ezért a DNS megkettőződésének folyamata eltérő, félig konzervatív módon: a DNS molekula feltekercselődik, és mindegyik láncon a komplementaritás elve szerint szintetizálódik egy új lánc. A DNS-molekula önreprodukciós folyamatát, amely biztosítja az örökletes információ pontos másolását és nemzedékről nemzedékre való továbbadását, ún. replikáció(a lat. replikáció- ismétlés). A replikáció eredményeként a kiindulási DNS-molekulából két abszolút pontos másolat keletkezik, amelyek mindegyike a szülő egy-egy példányát hordozza.

A replikáció folyamata valójában rendkívül összetett, mivel számos fehérje vesz részt benne. Egyesek a DNS kettős hélixét feltekerik, mások hidrogénkötéseket szakítanak meg a komplementer láncok nukleotidjai között, mások (például a DNS-polimeráz enzim) a komplementaritás elve alapján új nukleotidokat választanak ki stb. Két DNS-molekula keletkezik A replikáció kettévált az osztódás során.újonnan képződött leánysejtek.
A replikációs folyamat során fellépő hibák rendkívül ritkák, de ha mégis előfordulnak, akkor a DNS polimerázok és a speciális javító enzimek nagyon gyorsan kiküszöbölik őket, mivel a nukleotidszekvencia bármilyen hibája a fehérje szerkezetének és funkcióinak visszafordíthatatlan megváltozásához vezethet. és végső soron hátrányosan befolyásolják az életképességet új sejt vagy akár egyének.
fehérje bioszintézis. Ahogy a 19. század kiváló filozófusa, F. Engels képletesen fogalmazott: "Az élet a fehérjetestek létformája." A fehérjemolekulák szerkezetét és tulajdonságait elsődleges szerkezetük, azaz a DNS-ben kódolt aminosavak sorrendje határozza meg. Nemcsak magának a polipeptidnek a létezése, hanem a sejt egészének működése is függ ezen információ reprodukálásának pontosságától, ezért a fehérjeszintézis folyamata nagy jelentőséggel bír. Úgy tűnik, ez a legösszetettebb szintézisfolyamat a sejtben, mivel itt akár háromszáz különböző enzim és más makromolekula is részt vesz. Ráadásul nagy sebességgel folyik, ami még nagyobb pontosságot igényel.
A fehérje bioszintézisének két fő lépése van: a transzkripció és a transzláció.
Átírás(a lat. átírás- újraírás) az mRNS-molekulák bioszintézise DNS-templáton.

Mivel a DNS-molekula két antiparallel láncot tartalmaz, így mindkét láncból származó információ leolvasása teljesen eltérő mRNS-ek képződését eredményezné, ezért ezek bioszintézise csak az egyik láncon lehetséges, amit kódolónak vagy kodogénnek neveznek, ellentétben a másodikkal. nem kódoló vagy nem kodogén. Az újraírási folyamatot egy speciális enzim, az RNS-polimeráz biztosítja, amely a komplementaritás elve alapján választja ki az RNS-nukleotidokat. Ez a folyamat mind a sejtmagban, mind a saját DNS-sel rendelkező organellumokban - mitokondriumokban és plasztidokban - végbemehet.
A transzkripció során szintetizált mRNS-molekulák a transzlációhoz való komplex előkészítési folyamaton mennek keresztül (a mitokondriális és plasztid mRNS-ek organellumok belsejében maradhatnak, ahol a fehérje bioszintézis második szakasza zajlik). Az mRNS érési folyamata során az első három nukleotid (AUG) és egy adenil nukleotidok farka kapcsolódik hozzá, amelyek hossza határozza meg, hogy egy adott molekulán hány fehérje kópia szintetizálható. Az érett mRNS-ek csak ezután hagyják el a sejtmagot a magpórusokon keresztül.
Ezzel párhuzamosan a citoplazmában végbemegy az aminosav aktiválódási folyamata, melynek során az aminosav a megfelelő szabad tRNS-hez kötődik. Ezt a folyamatot egy speciális enzim katalizálja, ATP-t fogyaszt.
Adás(a lat. adás- transzfer) egy polipeptidlánc bioszintézise egy mRNS-templáton, amelyben a genetikai információ a polipeptidlánc aminosav-szekvenciájává alakul át.

A fehérjeszintézis második szakasza leggyakrabban a citoplazmában fordul elő, például a durva endoplazmatikus retikulumon. Előfordulásához riboszómák jelenléte, tRNS aktiválása, mely során a megfelelő aminosavakat megköti, Mg2+ ionok jelenléte, valamint optimális környezeti feltételek (hőmérséklet, pH, nyomás stb.) szükségesek.
A sugárzás megkezdéséhez megindítás, inicializálás) a riboszóma kis alegysége a szintézisre kész mRNS molekulához kapcsolódik, majd a komplementaritás elve szerint az első kodonhoz (AUG) szelektálják a metionin aminosavat hordozó tRNS-t. Csak ezután csatlakozik a riboszóma nagy alegysége. Az összeállított riboszómán belül két mRNS kodon található, amelyek közül az első már foglalt. A szomszédos kodonhoz egy második, szintén aminosavat hordozó tRNS kapcsolódik, majd az aminosavak között enzimek segítségével peptidkötés jön létre. A riboszóma az mRNS egy kodonját mozgatja; az aminosavtól megszabadított tRNS első része visszakerül a citoplazmába a következő aminosavért, a leendő polipeptidlánc egy fragmentuma pedig mintegy lóg a maradék tRNS-en. A következő tRNS csatlakozik az új kodonhoz, amely a riboszómán belül van, a folyamat megismétlődik és a polipeptid lánc lépésről lépésre meghosszabbodik, azaz megnyúlás.
A fehérjeszintézis vége megszüntetése) akkor következik be, amikor egy olyan mRNS-molekulában találkozunk egy adott nukleotidszekvenciával, amely nem kódol aminosavat (stopkodon). Ezt követően a riboszóma, az mRNS és a polipeptidlánc szétválik, majd az újonnan szintetizált fehérje megfelelő szerkezetet kap, és a sejtnek arra a részére kerül, ahol ellátja funkcióját.
A transzláció nagyon energiaigényes folyamat, mivel egy ATP-molekula energiáját egy aminosav tRNS-hez való kapcsolására fordítják, és több másikat használnak fel a riboszóma mozgatására az mRNS-molekula mentén.
Bizonyos fehérjemolekulák szintézisének felgyorsítása érdekében az mRNS-molekulához egymás után több riboszóma kapcsolódhat, amelyek egyetlen szerkezetet alkotnak - poliszóma.
A sejt az élőlények genetikai egysége. A kromoszómák, szerkezetük (alakjuk és méretük) és funkcióik. A kromoszómák száma és fajállandósága. Szomatikus és nemi sejtek. A sejt életciklusa: interfázis és mitózis. A mitózis a szomatikus sejtek osztódása. Meiosis. A mitózis és a meiózis fázisai. A csírasejtek fejlődése növényekben és állatokban. A sejtosztódás az élőlények növekedésének, fejlődésének és szaporodásának alapja. A meiózis és a mitózis szerepe
A sejt az élet genetikai egysége
Annak ellenére, hogy a nukleinsavak a genetikai információ hordozói, ezeknek az információknak a megvalósítása a sejten kívül lehetetlen, amit a vírusok példája is könnyen bizonyít. Ezek a gyakran csak DNS-t vagy RNS-t tartalmazó organizmusok nem képesek önmagukban szaporodni, ehhez a sejt örökletes apparátusát kell használniuk. Magának a sejtnek a segítsége nélkül még a sejtbe sem tudnak behatolni, kivéve a membrántranszport mechanizmusainak felhasználásával vagy sejtkárosodás következtében. A legtöbb vírus instabil, néhány óra szabad levegőn való tartózkodás után elpusztul. Ezért a sejt egy élőlény genetikai egysége, amely rendelkezik egy minimális összetevőkészlettel az örökletes információk megőrzéséhez, módosításához és megvalósításához, valamint az utódokhoz való továbbításához.
Az eukarióta sejt genetikai információinak nagy része a sejtmagban található. Szervezetének sajátossága, hogy a prokarióta sejt DNS-ével ellentétben az eukarióta DNS-molekulák nincsenek zárva, és komplex komplexeket képeznek a fehérjékkel - kromoszómákkal.
A kromoszómák, szerkezetük (alakjuk és méretük) és funkcióik
Kromoszóma(görögből. króm- szín, szín és harcsa- test) a sejtmag szerkezete, amely géneket tartalmaz, és bizonyos örökletes információkat hordoz a szervezet jeleiről és tulajdonságairól.
Néha a prokarióták gyűrűs DNS-molekuláit kromoszómáknak is nevezik. A kromoszómák képesek önmegkettőzésre, szerkezeti és funkcionális egyéniségük van és azt több generáción keresztül megőrzik. Minden sejt hordozza a test összes örökletes információját, de csak egy kis része működik.
A kromoszóma alapja egy fehérjékkel teli, kétszálú DNS-molekula. Az eukariótákban a hiszton és a nem hiszton fehérjék kölcsönhatásba lépnek a DNS-sel, míg a prokariótákban a hiszton fehérjék hiányoznak.
A kromoszómák fénymikroszkóp alatt a sejtosztódás során láthatók legjobban, amikor a tömörítés következtében rúd alakú testek formáját öltik, amelyeket elsődleges szűkület választ el egymástól - centromer — vállán. A kromoszómának is lehet másodlagos szűkület, amely bizonyos esetekben elválasztja az ún műhold. A kromoszómák végeit ún telomerek. A telomerek megakadályozzák, hogy a kromoszómák végei összetapadjanak, és biztosítják a nukleáris membránhoz való kapcsolódásukat egy nem osztódó sejtben. Az osztódás kezdetén a kromoszómák megduplázódnak, és két leánykromoszómából állnak - kromatidák a centromérához rögzítve.

Az alak szerint egyenlő karú, egyenlőtlen karú és rúd alakú kromoszómákat különböztetünk meg. A kromoszómák mérete jelentősen eltér, de az átlagos kromoszóma mérete 5 $ × $ 1,4 µm.
Egyes esetekben a kromoszómák számos DNS-duplikáció eredményeként több száz és ezer kromatidot tartalmaznak: az ilyen óriáskromoszómákat ún. polietilén. Találkoznak nyálmirigyek Drosophila lárvák, valamint az orsóférgek emésztőmirigyeiben.
A kromoszómák száma és fajállandósága. Szomatikus és csírasejtek
A sejtelmélet szerint a sejt egy szervezet szerkezetének, életének és fejlődésének egysége. Így az élőlények olyan fontos funkcióit, mint a növekedés, a szaporodás és a szervezet fejlődése sejtszinten biztosítják. A többsejtű szervezetek sejtjei szomatikus és nemi csoportokra oszthatók.
szomatikus sejtek a test összes sejtje, amely mitotikus osztódás eredményeként képződik.
A kromoszómák vizsgálata lehetővé tette annak megállapítását, hogy az egyes biológiai fajok szervezetének szomatikus sejtjeit állandó számú kromoszóma jellemzi. Például egy embernek 46. A szomatikus sejtek kromoszómáinak halmazát ún diploid(2n), vagy dupla.

nemi sejtek, vagy ivarsejtek, speciális sejtek, amelyek az ivaros szaporodást szolgálják.
Az ivarsejtek mindig feleannyi kromoszómát tartalmaznak, mint a szomatikus sejtekben (emberben - 23), ezért a csírasejtek kromoszómáinak halmazát ún. haploid(n), vagy egyetlen. Kialakulása a meiotikus sejtosztódáshoz kapcsolódik.
A szomatikus sejtek DNS mennyisége 2c, a csírasejteké pedig 1c. A szomatikus sejtek genetikai képlete 2n2c, a neme pedig 1n1c.
Egyes szomatikus sejtek magjában a kromoszómák száma eltérhet a szomatikus sejtek számától. Ha ez a különbség egy, kettő, három stb. haploid halmazzal nagyobb, akkor az ilyen sejteket hívjuk poliploid(tri-, tetra-, pentaploid, ill. Az ilyen sejtekben az anyagcsere folyamatok általában nagyon intenzívek.
A kromoszómák száma önmagában nem fajspecifikus tulajdonság, hiszen a különböző organizmusok azonos számú kromoszómával rendelkezhetnek, míg a rokon élőlények eltérő számmal rendelkezhetnek. Például a maláriás plazmódiumnak és a ló orsóféregnek két kromoszómája van, míg az embernek és a csimpánznak 46, illetve 48 kromoszómája.
Az emberi kromoszómákat két csoportra osztják: autoszómákra és nemi kromoszómákra (heterokromoszómák). Autosome az emberi szomatikus sejtekben 22 pár van, férfiaknál és nőknél azonosak, ill nemi kromoszómák csak egy pár, de ő határozza meg az egyed nemét. Kétféle nemi kromoszóma létezik - X és Y. A nők testének sejtjei két X kromoszómát hordoznak, a férfiak pedig X és Y kromoszómát.
Kariotípus- ez egy szervezet kromoszómakészletének jeleinek halmaza (a kromoszómák száma, alakja és mérete).
A kariotípus feltételes rekordja tartalmazza a kromoszómák teljes számát, a nemi kromoszómákat és a kromoszómakészlet lehetséges eltéréseit. Például egy normál férfi kariotípusa 46,XY, míg egy normál nő kariotípusa 46,XX.
A sejt életciklusa: interfázis és mitózis
A sejtek nem keletkeznek minden alkalommal újra, csak az anyasejtek osztódása következtében jönnek létre. Az elválasztást követően a leánysejteknek némi időbe telik, hogy organellumokat képezzenek, és megszerezzék a megfelelő szerkezetet, amely biztosítaná egy bizonyos funkció ellátását. Ezt az időszakot ún érés.
A sejt osztódás eredményeként való megjelenésétől az osztódásig vagy haláláig eltelt időt nevezzük sejt életciklusa.
Az eukarióta sejtekben az életciklus két fő szakaszra oszlik: interfázisra és mitózisra.
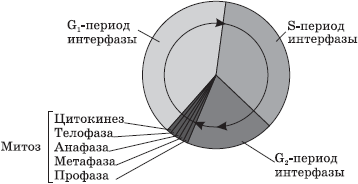
Interfázis- ez az az időszak az életciklusban, amikor a sejt nem osztódik és normálisan működik. Az interfázis három szakaszra oszlik: G 1 -, S- és G 2 - periódusra.
G 1 -periódus(preszintetikus, posztmitotikus) a sejtnövekedés és fejlődés időszaka, mely során az újonnan képződött sejt teljes életfenntartásához szükséges RNS, fehérjék és egyéb anyagok aktív szintézise zajlik. Ezen időszak végére a sejt elkezdhet felkészülni a DNS-megkettőzésre.
NÁL NÉL S-időszak(szintetikus) a DNS-replikáció folyamata megy végbe. A kromoszóma egyetlen része, amely nem replikálódik, a centromer, ezért a létrejövő DNS-molekulák nem válnak szét teljesen, hanem rögzítve maradnak benne, és az osztódás kezdetén a kromoszóma X-alakú megjelenésű. A sejt genetikai képlete a DNS megkettőződése után 2n4c. Szintén az S-periódusban a sejtközpont centrioljainak megkettőződése következik be.
G 2 -periódus(posztszintetikus, premitotikus) a sejtosztódási folyamathoz szükséges RNS, fehérjék és ATP intenzív szintézise, valamint a centriolok, mitokondriumok és plasztidok szétválása jellemzi. Az interfázis végéig a kromatin és a nucleolus jól elkülöníthető marad, a magmembrán integritása nem sérül, az organellumok nem változnak.
A test sejtjeinek egy része a szervezet egész életében képes ellátni funkcióit (agyunk idegsejtjei, szív izomsejtjei), míg mások rövid ideig léteznek, majd elpusztulnak (a bélhám sejtjei). , a bőr hámrétegének sejtjei). Következésképpen a szervezetben folyamatosan zajlanak a sejtosztódási folyamatok, új sejtek képződése, amelyek az elhaltak helyébe lépnek. Az osztódásra képes sejteket ún származik. Az emberi testben piros színűek csontvelő, a bőr hámrétegének mély rétegeiben és más helyeken. Ezekkel a sejtekkel új szervet növeszthet, megfiatalíthat, és klónozhatja a testet. Az őssejtek felhasználásának kilátásai meglehetősen világosak, de ennek a problémának a morális és etikai vonatkozásait még vitatják, mivel a legtöbb esetben abortusz során elpusztult emberi magzatokból nyert embrionális őssejteket használnak.
A növényi és állati sejtekben az interfázis időtartama átlagosan 10-20 óra, míg a mitózis körülbelül 1-2 órát vesz igénybe.
A többsejtű élőlényekben az egymást követő osztódások során a leánysejtek egyre változatosabbá válnak, mivel egyre több génről olvasnak információt.
Egyes sejtek végül abbahagyják az osztódást és elpusztulnak, ami bizonyos funkciók – például a bőr hámsejtjei és a vérsejtek esetében – vagy e sejtek környezeti tényezők, különösen kórokozók általi károsítása miatt következhet be. A genetikailag programozott sejthalált nevezik apoptózis míg a véletlen halál az elhalás.
A mitózis a szomatikus sejtek osztódása. A mitózis fázisai
Mitózis- a szomatikus sejtek közvetett osztódásának módszere.
A mitózis során a sejt egy sor egymást követő fázison megy keresztül, melynek eredményeként minden leánysejt ugyanazt a kromoszómakészletet kapja, mint az anyasejtben.
A mitózis négy fő fázisra oszlik: profázisra, metafázisra, anafázisra és telofázisra. Prophase- a mitózis leghosszabb szakasza, melynek során kromatinkondenzáció megy végbe, melynek eredményeként két kromatidából (leánykromoszómák) álló X alakú kromoszómák válnak láthatóvá. Ebben az esetben a nucleolus eltűnik, a centriolák a sejt pólusai felé eltérnek, és elkezd kialakulni a mikrotubulusok akromatin orsója (orsója). A profázis végén a magmembrán különálló vezikulákra bomlik.

NÁL NÉL metafázis a kromoszómák a sejt egyenlítője mentén sorakoznak centromereikkel, amelyekhez egy teljesen kialakult osztódási orsó mikrotubulusai kapcsolódnak. Az osztódásnak ebben a szakaszában a kromoszómák a legsűrűbbek és jellegzetes formájúak, ami lehetővé teszi a kariotípus tanulmányozását.
NÁL NÉL anafázis A centromerekben gyors DNS-replikáció megy végbe, melynek eredményeként a kromoszómák felhasadnak, és a kromatidák a sejt mikrotubulusokkal megfeszített pólusai felé eltérnek. A kromatidák eloszlásának abszolút egyenlőnek kell lennie, mivel ez a folyamat tartja fenn a kromoszómák számának állandóságát a test sejtjeiben.
A színpadon telofázis a leánykromoszómák a pólusokon összegyűlnek, despiralizálódnak, körülöttük a vezikulákból magburok képződik, az újonnan képződött magvakban pedig sejtmagok jelennek meg.
A sejtmag osztódása után megtörténik a citoplazma osztódása - citokinézis, amely során többé-kevésbé egyenletesen oszlik el az anyasejt összes organellumja.
Így a mitózis hatására egy anyasejtből két leánysejt képződik, amelyek mindegyike az anyasejt genetikai másolata (2n2c).
A beteg, sérült, öregedő sejtekben és a test speciális szöveteiben egy kissé eltérő osztódási folyamat - amitózis - léphet fel. Amitózis az eukarióta sejtek közvetlen osztódásának nevezik, amelyben a genetikailag egyenértékű sejtek képződése nem történik meg, mivel a sejtkomponensek egyenetlenül oszlanak el. Növényekben az endospermiumban, állatokban pedig a májban, a porcokban és a szem szaruhártyájában fordul elő.
Meiosis. A meiózis fázisai
Meiosis- ez az elsődleges csírasejtek (2n2c) közvetett osztódásának módszere, melynek eredményeként haploid sejtek (1n1c), leggyakrabban csírasejtek képződnek.
A mitózistól eltérően a meiózis két egymást követő sejtosztódásból áll, mindegyiket egy interfázis előzi meg. A meiózis első felosztását (meiosis I.) ún csökkentés, mivel ebben az esetben a kromoszómák száma felére csökken, és a második osztódás (meiosis II) - egyenlítő, mivel ennek során a kromoszómák száma megmarad.

I. interfázis hasonlóan halad a mitózis interfázisához. Meiosis I négy fázisra oszlik: I. profázis, I. metafázis, I. anafázis és I. telofázis. Prófázis I Két fő folyamat játszódik le: a konjugáció és a keresztezés. Konjugáció- ez a homológ (páros) kromoszómák teljes hosszában történő fúziójának folyamata. A konjugáció során keletkezett kromoszómapárok az I. metafázis végéig megmaradnak.
Átkelés- a homológ kromoszómák homológ régióinak kölcsönös cseréje. A keresztezés eredményeként a szervezet által mindkét szülőtől kapott kromoszómák új génkombinációkat szereznek, ami genetikailag változatos utódok megjelenéséhez vezet. Az I. profázis végén, akárcsak a mitózis profázisában, a sejtmag eltűnik, a centriolák a sejt pólusai felé eltérnek, és a magburok felbomlik.

NÁL NÉL metafázis I a sejt egyenlítője mentén kromoszómapárok sorakoznak fel, centromereikhez a hasadási orsó mikrotubulusai kapcsolódnak.
NÁL NÉL anafázis I két kromatidából álló egész homológ kromoszómák eltérnek a pólusokhoz.
NÁL NÉL telofázis I a sejt pólusain lévő kromoszómacsoportok körül magmembránok, sejtmagvak alakulnak ki.
Citokinézis I biztosítja a leánysejtek citoplazmájának osztódását.
A meiózis I (1n2c) eredményeként létrejövő leánysejtek genetikailag heterogének, mivel a sejt pólusaira véletlenszerűen szétszórt kromoszómáik nem egyenlő géneket tartalmaznak.
A mitózis és a meiózis összehasonlító jellemzői
| jel | Mitózis | Meiosis | |
| Milyen sejtek kezdenek osztódni? | Szomatikus (2n) | Elsődleges csírasejtek (2n) | |
| Osztályok száma | 1 | 2 | |
| Hány és milyen sejt képződik az osztódás során? | 2 szomatikus (2n) | 4 szexuális (n) | |
| Interfázis | A sejtek felkészítése osztódásra, DNS megkettőzésre | Nagyon rövid, a DNS megkettőződése nem fordul elő | |
| Fázisok | Meiosis I | Meiosis II | |
| Prophase | Előfordulhat kromoszóma kondenzáció, a sejtmag eltűnése, a magburok szétesése, konjugáció és keresztezés | A kromoszómák kondenzációja, a sejtmag eltűnése, a magburok szétesése | |
| metafázis | Az egyenlítő mentén kromoszómapárok helyezkednek el, osztódási orsó képződik | A kromoszómák az Egyenlítő mentén sorakoznak, kialakul az osztódási orsó | |
| Anafázis | A két kromatidából származó homológ kromoszómák a pólusok felé eltérnek | A kromatidák a pólusok felé eltérnek | |
| Telofázis | A kromoszómák despiralizálódnak, új magburok és nukleolusok képződnek | A kromoszómák despiralizálódnak, új magburok és nukleolusok képződnek | |
Interfázis II nagyon rövid, mivel nem történik meg benne a DNS megkettőződése, vagyis nincs S-periódus.
Meiosis II szintén négy fázisra oszlik: II. profázis, II. metafázis, II. anafázis és II. telofázis. NÁL NÉL prófázis II ugyanazok a folyamatok mennek végbe, mint az I. prózában, kivéve a ragozást és az átkelést.
NÁL NÉL metafázis II A kromoszómák a sejt egyenlítője mentén helyezkednek el.
NÁL NÉL anafázis II A kromoszómák a centromérán hasadnak, és a kromatidák a pólusok felé nyúlnak.
NÁL NÉL telofázis II nukleáris membránok és nukleolusok képződnek a leánykromoszómák klaszterei körül.
Után citokinézis II mind a négy leánysejt genetikai képlete 1n1c, de mindegyiknek más a génkészlete, ami az anyai és apai kromoszómák keresztezésének és véletlenszerű kombinációjának az eredménye a leánysejtekben.
A csírasejtek fejlődése növényekben és állatokban
Gametogenezis(görögből. ivarsejt- feleség, ivarsejtek- férj és keletkezése- eredet, előfordulás) az érett csírasejtek képződésének folyamata.
Mivel az ivaros szaporodáshoz leggyakrabban két egyedre van szükség - nőstényre és hímre, akik különböző nemi sejteket - petesejteket és spermiumokat termelnek, akkor ezeknek az ivarsejteknek a képződési folyamatainak eltérőnek kell lenniük.
A folyamat jellege nagymértékben függ attól is, hogy növényi vagy állati sejtben történik-e, hiszen a növényekben az ivarsejtek képződése során csak a mitózis, míg az állatoknál a mitózis és a meiózis egyaránt előfordul.
A csírasejtek fejlődése a növényekben. A zárvatermőkben a hím és nőstény csírasejtek képződése a virág különböző részein - porzókban, illetve bibékben - történik.
A férfi csírasejtek kialakulása előtt - mikrogametogenezis(görögből. mikros- kicsi) - történik mikrosporogenezis, vagyis a porzók porzójában a mikrospórák képződése. Ez a folyamat az anyasejt meiotikus osztódásához kapcsolódik, melynek eredményeként négy haploid mikrospóra keletkezik. A mikrogametogenezis a mikrospórák mitotikus osztódásával jár, ami két sejtből álló hím gametofitot eredményez - egy nagy vegetatív(szifonogén) és sekély generatív. Az osztódás után a hím gametofitot sűrű héjak borítják, és pollenszemcsét képeznek. Egyes esetekben a generatív sejt még a pollenérés folyamatában is, néha csak a bibe stigmájába történő áthelyezés után mitotikusan osztódik, két mozdulatlan hím csírasejt képződésével - sperma. A beporzás után a vegetatív sejtből pollencső képződik, amelyen keresztül a spermiumok behatolnak a bibe petefészkébe a megtermékenyítés céljából.

A női csírasejtek fejlődését a növényekben ún megagametogenezis(görögből. megas- nagy). A bibe petefészkében fordul elő, amelyet megelőz megasporogenezis, melynek eredményeként a nucellusban fekvő megaspóra anyasejtjéből meiotikus osztódással négy megaspóra képződik. Az egyik megaspóra mitotikusan háromszor osztódik, és létrejön a női gametofiton, egy nyolc maggal rendelkező embriózsák. A leánysejtek citoplazmájának ezt követő izolálásával az egyik létrejövő sejt tojássá válik, melynek oldalain az úgynevezett szinergiák találhatók, az embriózsák másik végén három antipód képződik, középen pedig , két haploid mag fúziója eredményeként diploid központi sejt keletkezik.
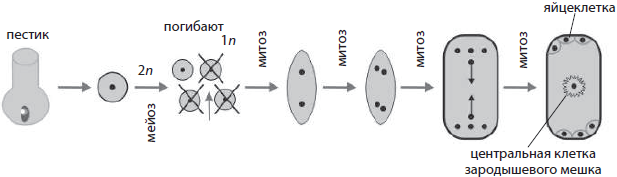
A csírasejtek fejlődése állatokban. Az állatokban a csírasejtek képződésének két folyamata különböztethető meg - a spermatogenezis és az oogenezis.
spermatogenezis(görögből. spermiumok, spermiumok- vetőmag és keletkezése- eredet, előfordulás) az érett hím csírasejtek - spermiumok képződésének folyamata. Emberben a herékben vagy herékben fordul elő, és négy szakaszra oszlik: szaporodás, növekedés, érés és képződés.

NÁL NÉL tenyésztési időszak a primordiális csírasejtek mitotikusan osztódnak, ami diploid képződést eredményez spermatogónia. NÁL NÉL növekedési időszak A spermatogóniák tápanyagokat halmoznak fel a citoplazmában, megnövekednek és átalakulnak elsődleges spermatociták, vagy I. rendű spermatociták. Csak ezután lépnek be a meiózisba ( érési időszak), ami először kettőt eredményez másodlagos spermatocita, vagy 2. rendű spermatociták, majd - négy haploid sejt meglehetősen nagy mennyiségű citoplazmával - spermatidok. NÁL NÉL kialakulásának időszaka szinte az egész citoplazmát elveszítik, és flagellumot alkotnak, amely spermiumokká alakul.
spermiumok, vagy gumicukorkák, - nagyon kicsi, mozgékony férfi nemi sejtek fejjel, nyakkal és farokkal.

NÁL NÉL fej, kivéve a magot, van akroszóma- módosított Golgi komplex, amely biztosítja a petesejt membránjainak feloldódását a megtermékenyítés során. NÁL NÉL nyak ott vannak a sejtközpont centrioljai, és az alap lófarok mikrotubulusokat képeznek, amelyek közvetlenül támogatják a spermium mozgását. Mitokondriumokat is tartalmaz, amelyek a spermiumot ATP energiával látják el a mozgáshoz.
Ovogenezis(görögből. ENSZ- egy tojás és keletkezése- eredet, előfordulás) az érett női csírasejtek - peték kialakulásának folyamata. Emberben a petefészkekben fordul elő, és három szakaszból áll: szaporodás, növekedés és érés. A spermatogenezishez hasonló szaporodási és növekedési periódusok még az intrauterin fejlődés során is előfordulnak. Ugyanakkor az elsődleges csírasejtekből a mitózis hatására diploid sejtek képződnek. oogonia, amelyek aztán diploid elsődlegesvé alakulnak petesejtek, vagy I. rendű petesejtek. A meiózis és az azt követő citokinézis a érési időszak, az anyasejt citoplazmájának egyenetlen osztódása jellemzi, így ennek eredményeként először egy másodlagos petesejt, vagy petesejt 2. rend, és első poláris test, majd a másodlagos petesejtből - a tojásból, amely megtartja a teljes tápanyagellátást, és a második poláris testet, míg az első poláris test két részre oszlik. A sarki testek elveszik a felesleges genetikai anyagot.
Emberben a tojások 28-29 napos időközönként keletkeznek. A tojások érésével és felszabadulásával kapcsolatos ciklust menstruációs ciklusnak nevezik.
Tojás- egy nagy női ivarsejt, amely nemcsak haploid kromoszómakészletet hordoz, hanem jelentős tápanyagellátást is biztosít az embrió későbbi fejlődéséhez.

Az emlősök tojását négy membrán borítja, amelyek csökkentik a különböző tényezők által okozott károsodás valószínűségét. A tojás átmérője emberben eléri a 150-200 mikront, míg a struccnál több centiméter is lehet.
A sejtosztódás az élőlények növekedésének, fejlődésének és szaporodásának alapja. A mitózis és a meiózis szerepe
Ha az egysejtű szervezetekben a sejtosztódás az egyedek számának növekedéséhez, azaz a szaporodáshoz vezet, akkor a többsejtű szervezetekben ennek a folyamatnak más jelentése is lehet. Így az embrió sejtosztódása a zigótától kezdve a biológiai alapja a növekedés és fejlődés egymással összefüggő folyamatainak. Hasonló változások figyelhetők meg az emberekben is serdülőkor amikor nemcsak a sejtek száma nő, hanem minőségi változás is bekövetkezik a szervezetben. A többsejtű élőlények szaporodása is sejtosztódáson alapul, például az ivartalan szaporodás során ennek a folyamatnak köszönhetően egy testrészből az egész test helyreáll, az ivaros szaporodás során pedig a gametogenezis során csírasejtek keletkeznek, ezt követően adva új szervezet. Meg kell jegyezni, hogy az eukarióta sejtosztódás fő módszerei – a mitózis és a meiózis – eltérő jelentőséggel bírnak az élőlények életciklusában.
A mitózis következtében az örökítőanyag egyenletesen oszlik el a leánysejtek között - az anya pontos másolatai. Mitózis nélkül lehetetlen lenne egyetlen sejtből, a zigótából fejlődő többsejtű szervezetek létezése és növekedése, mivel az ilyen szervezetek minden sejtjének ugyanazt a genetikai információt kell tartalmaznia.
Az osztódás során a leánysejtek szerkezetükben és funkciójukban is egyre változatosabbá válnak, ami az intercelluláris kölcsönhatás következtében új géncsoportok aktiválódásával jár bennük. Tehát a mitózis szükséges a szervezet fejlődéséhez.
Ez a sejtosztódási módszer szükséges az ivartalan szaporodáshoz és a sérült szövetek, valamint szervek regenerációjához (helyreállításához).
A meiózis pedig biztosítja a kariotípus állandóságát az ivaros szaporodás során, mivel felére csökkenti az ivaros szaporodás előtti kromoszómakészletet, amely aztán a megtermékenyítés hatására helyreáll. Ezenkívül a meiózis a szülői gének új kombinációinak megjelenéséhez vezet a kromoszómák keresztezése és véletlenszerű kombinációja miatt a leánysejtekben. Ennek köszönhetően az utódok genetikailag változatosak, ami a természetes szelekció anyagát adja és az evolúció anyagi alapja. A kromoszómák számának, alakjának és méretének változása egyrészt a szervezet fejlődésében különböző eltérések megjelenéséhez, sőt akár halálához is vezethet, másrészt egyedek megjelenéséhez vezethet. jobban alkalmazkodnak a környezethez.
Így a sejt az élőlények növekedésének, fejlődésének és szaporodásának egysége.
A legtöbb élő szervezet sejtszerkezettel rendelkezik. A sejt az élővilág szerkezeti és funkcionális egysége. Az élő szervezetek összes jele és funkciója jellemzi: anyagcsere és energia, növekedés, szaporodás, önszabályozás. A sejtek formájukban, méretükben, funkciójukban, anyagcsere-típusukban különböznek egymástól (47. ábra).
Rizs. 47. Változatos sejtek: 1 - zöld euglena; 2 - baktérium; 3 - levélpép növényi sejtje; 4 - hámsejt; 5 - idegsejt
A sejtek mérete 3-10 és 100 µm között változik (1 µm = 0,001 m). Az 1-3 mikronnál kisebb sejtek ritkábban találkoznak. Vannak óriási sejtek is, amelyek mérete eléri a több centimétert. A sejtek alakja is nagyon változatos: gömb alakú, hengeres, ovális, orsó alakú, csillag alakú stb. Azonban sok a közös minden sejt között. Kémiai összetételük és általános szerkezeti tervük megegyezik.
A sejt kémiai összetétele. Az élő szervezetekben található összes ismert kémiai elem közül körülbelül 20 van, és közülük 4 részaránya: oxigén, szén, hidrogén és nitrogén - akár 95% -ot tesz ki. Ezeket az elemeket biogén elemeknek nevezzük. Az élő szervezeteket alkotó szervetlen anyagok közül a víz a legfontosabb. Tartalma a sejtben 60 és 98% között mozog. A sejt a víz mellett ásványi anyagokat is tartalmaz, főleg ionok formájában. Ezek a vas, jód, klór, foszfor, kalcium, nátrium, kálium stb. vegyületei.
A szervetlen anyagokon kívül szerves anyagok is jelen vannak a sejtben: fehérjék, lipidek (zsírok), szénhidrátok (cukrok), nukleinsavak (DNS, RNS). Ők teszik ki a sejt nagy részét. A legfontosabb szerves anyagok a nukleinsavak és a fehérjék. A nukleinsavak (DNS és RNS) részt vesznek az örökletes információk továbbításában, a fehérjeszintézisben, a sejt összes életfolyamatának szabályozásában.
Mókusok számos funkciót lát el: építési, szabályozási, szállítási, összehúzó, védelmi, energetikai. De a legfontosabb a fehérjék enzimatikus funkciója.
Enzimek- Ezek olyan biológiai katalizátorok, amelyek felgyorsítják és szabályozzák az élő szervezetekben végbemenő kémiai reakciók széles skáláját. Egy élő sejtben egyetlen reakció sem megy végbe enzimek részvétele nélkül.
Lipidekés szénhidrátokat főként építő és energetikai funkciókat látnak el, a szervezet tartalék tápanyagai.
Így, foszfolipidek A fehérjékkel együtt felépítik a sejt összes membránszerkezetét. Nagy molekulatömegű szénhidrát – a cellulóz a növények és gombák sejtfalát képezi.
Zsírok, keményítőés glikogén tartalék tápanyagok a sejt és a szervezet egésze számára. Glükóz, fruktóz, szacharóz és mások Szahara a növények gyökereinek és leveleinek, gyümölcseinek részei. Szőlőcukor az emberek és számos állat vérplazmájának elengedhetetlen alkotóeleme. A szénhidrátok és zsírok lebontásakor a szervezetben nagy mennyiségű energia szabadul fel, ami a létfontosságú folyamatokhoz szükséges.
Sejtszerkezetek. A sejt egy külső sejtmembránból, sejtszerveket tartalmazó citoplazmából és egy sejtmagból áll (48. ábra).

Rizs. 48. Egy állati (A) és növényi (B) sejt felépítésének kombinált sémája: 1 - héj; 2 - külső sejtmembrán 3 - mag; 4 - kromatin; 5 - nucleolus; 6 - endoplazmatikus retikulum (sima és szemcsés); 7 - mitokondriumok; 8 - kloroplasztiszok; 9 - Golgi készülék; 10 - lizoszóma; 11 - sejtközpont; 12 - riboszómák; 13 - vakuólum; 14 - citoplazma
külső sejtmembrán egy membránból álló sejtszerkezet, amely korlátozza minden élőlény sejtjének élő tartalmát. Szelektív permeabilitással rendelkezik, védi a sejtet, szabályozza az anyagáramlást és a külső környezettel való cserét, megtartja a sejt bizonyos formáját. Sejtek növényi szervezetek, gombák, a külső hártyán kívül héjuk is van. Ez az élettelen sejtszerkezet a növényekben cellulózból és a gombákban kitinből áll, erőt ad a sejtnek, védi azt, valamint a növények és gombák "csontváza".
NÁL NÉL citoplazma, a sejt félig folyékony tartalma, mind organellumok.
Endoplazmatikus retikulum behatol a citoplazmába, kommunikációt biztosítva a sejt egyes részei között és az anyagok szállítását. Vannak sima és szemcsés EPS-ek. A szemcsés ER riboszómákat tartalmaz.
Riboszómák- Ezek kis gomba alakú testek, amelyeken a sejtben fehérjeszintézis megy végbe.
golgi készülék biztosítja a szintetizált anyagok csomagolását és eltávolítását a sejtből. Ezenkívül struktúráiból alakulnak ki lizoszómák. Ezek a gömb alakú testek enzimeket tartalmaznak, amelyek lebontják a sejtbe jutó tápanyagokat, lehetővé téve az intracelluláris emésztést.
Mitokondriumok- Ezek hosszúkás alakú, félig autonóm membránszerkezetek. Számuk a sejtekben eltérő és az osztódás következtében növekszik. A mitokondriumok a sejt erőművei. A légzés során az anyagok végső oxidációja a légköri oxigénnel történik bennük. Ebben az esetben a felszabaduló energiát ATP molekulák tárolják, amelyek szintézise ezekben a struktúrákban történik.
kloroplasztiszok, félig autonóm membránszervecskék, amelyek csak a növényi sejtekre jellemzőek. A kloroplasztiszok zöld színűek a klorofill pigment miatt, ezek biztosítják a fotoszintézis folyamatát.
A kloroplasztokon kívül növényi sejtek van és vakuolák sejtnedvvel telve.
Cell Center részt vesz a sejtosztódás folyamatában. Két centriolból és egy centroszférából áll. Az osztódás során ezek alkotják a hasadási orsószálakat, és biztosítják a kromoszómák egyenletes eloszlását a sejtben.
Mag a sejtaktivitás szabályozásának központja. A sejtmagot a citoplazmától egy magmembrán választja el, amelynek pórusai vannak. Belseje tele van karioplazmával, amely olyan DNS-molekulákat tartalmaz, amelyek biztosítják az örökletes információk továbbítását. Itt történik a DNS, RNS, riboszómák szintézise. Gyakran egy vagy több sötét, lekerekített képződmény látható a sejtmagban - ezek a nucleolusok. Itt riboszómák képződnek és halmozódnak fel. A sejtmagban a DNS-molekulák nem láthatók, mivel vékony kromatinszálak formájában vannak. Az osztódás előtt a DNS spiralizálódik, megvastagodik, komplexeket képez a fehérjével, és jól látható struktúrákká - kromoszómákká - alakul (49. ábra). Általában a sejtben lévő kromoszómák párosak, alakjuk, méretük és örökletes információjuk azonos. A páros kromoszómákat ún homológ. A kromoszómák kettős halmazát nevezzük diploid. Egyes sejtek és organizmusok egyetlen, párosítatlan halmazt tartalmaznak, az úgynevezett haploid.

Rizs. 49. A - a kromoszóma szerkezete: 1 - centromer; 2 - kromoszóma karok; 3 - DNS-molekulák; 4 - testvérkromatidák B - a kromoszómák típusai: 1 - egyenlő vállúak; 2 - több vállú; 3 - egyetlen váll
A kromoszómák száma minden szervezettípushoz állandó. Így az emberi sejtekben 46 kromoszóma (23 pár), a búzasejtekben 28 (14 pár), a galambsejtekben 80 (40 pár) található. Ezek a szervezetek diploid kromoszómákat tartalmaznak. Egyes organizmusok, például algák, mohák, gombák haploid kromoszómakészlettel rendelkeznek. A nemi sejtek minden szervezetben haploidok.
A felsoroltakon kívül néhány sejtnek specifikus organellumja is van - csillókés flagella, mozgást biztosítanak elsősorban az egysejtű szervezetekben, de jelen vannak a többsejtű szervezetek egyes sejtjeiben is. Például a flagellák a zöld euglena-ban, a chlamydomonas-ban és néhány baktériumban, a csillók pedig a csillószájban, az állatok csillóhámsejtjeiben találhatók.
| |
43. § Az életvitel fő kritériumai45. § A sejt élettevékenységének jellemzői
Hasonló oldalak
Az élő szervezetek kémiai összetétele
Az élő szervezetek kémiai összetétele két formában fejezhető ki: atomi és molekuláris formában. Az atomi (elemi) összetétel az élő szervezeteket alkotó elemek atomjainak arányát mutatja. A molekuláris (anyag) összetétel az anyagok molekuláinak arányát tükrözi.
A kémiai elemek a sejtek részei ionok és szervetlen és szerves anyagok molekulái formájában. A sejtben a legfontosabb szervetlen anyagok a víz és az ásványi sók, a legfontosabb szerves anyagok a szénhidrátok, lipidek, fehérjék és nukleinsavak.
A víz minden élő szervezet domináns alkotóeleme. A legtöbb élő szervezet sejtjeinek átlagos víztartalma körülbelül 70%.
Az ásványi sók a sejt vizes oldatában kationokká és anionokká disszociálnak. A legfontosabb kationok a K+, Ca2+, Mg2+, Na+, NHJ, anionok - Cl-, SO2-, HPO2-, H2PO-, HCO-, NO-.
Szénhidrát - szerves vegyületek, amelyek egy vagy több egyszerű cukormolekulából állnak. Az állati sejtekben a szénhidráttartalom 1-5%, egyes növényi sejtekben pedig eléri a 70%-ot.
Lipidek - zsírok és zsírszerű szerves vegyületek, vízben gyakorlatilag nem oldódnak. Tartalmuk a különböző sejtekben nagyon változó: 2-3-tól 50-90%-ig a növényi magvak sejtjeiben és az állatok zsírszövetében.
Mókusok olyan biológiai heteropolimerek, amelyek monomerjei aminosavak. Csak 20 aminosav vesz részt a fehérjék képződésében. Ezeket alapvetőnek vagy alapvetőnek nevezik. Az aminosavak egy része nem szintetizálódik az állatok és az emberek szervezetében, ezért növényi táplálékkal kell ellátni őket (ezeket esszenciálisnak nevezik).
Nukleinsavak. Kétféle nukleinsav létezik: DNS és RNS. A nukleinsavak olyan polimerek, amelyek monomerjei nukleotidok.
Sejtszerkezet
A sejtelmélet kialakulása
- Robert Hooke 1665-ben sejteket fedezett fel egy parafa metszetben, és ő használta először a "sejt" kifejezést.
- Anthony van Leeuwenhoek egysejtű szervezeteket fedezett fel.
- Matthias Schleiden 1838-ban és Thomas Schwann 1839-ben fogalmazta meg a sejtelmélet főbb rendelkezéseit. Azonban tévesen azt hitték, hogy a sejtek az elsődleges, nem sejtes anyagból származnak.
- Rudolf Virchow 1858-ban bebizonyította, hogy minden sejt más sejtekből sejtosztódással jön létre.
A sejtelmélet alapvető rendelkezései
- A sejt minden élőlény szerkezeti egysége. Minden élő szervezet sejtekből áll (a vírusok kivételt képeznek).
- A sejt minden élőlény funkcionális egysége. A sejt a létfontosságú funkciók teljes skáláját mutatja.
- A sejt minden élőlény fejlődési egysége. Új sejtek csak az eredeti (anya) sejt osztódása következtében jönnek létre.
- A sejt minden élőlény genetikai egysége. A sejt kromoszómái az egész szervezet fejlődéséről tartalmaznak információkat.
- Minden élőlény sejtje hasonló kémiai összetételben, szerkezetben és működésben.
A sejtszerveződés típusai
Az élő szervezetek közül csak a vírusok nem rendelkeznek sejtszerkezettel. Az összes többi organizmust sejtes életformák képviselik. A sejtszerveződésnek két típusa van: prokarióta és eukarióta. A baktériumok prokarióták, a növények, gombák és állatok pedig eukarióták.
A prokarióta sejtek viszonylag egyszerűek. Magjuk nincs, a citoplazmában a DNS helyét nukleoidnak nevezik, az egyetlen DNS molekula kör alakú és nem kapcsolódik fehérjékhez, a sejtek kisebbek, mint az eukarióta sejtek, a sejtfal glikopeptidet - mureint tartalmaz, vannak membránszervecskék nincsenek, funkciójukat a plazmamembrán invaginációi látják el, a riboszómák kicsik, a mikrotubulusok hiányoznak, így a citoplazma mozdulatlan, a csillók és flagellák speciális szerkezetűek.
Az eukarióta sejteknek van egy magja, amelyben a kromoszómák találhatók - fehérjékhez kapcsolódó lineáris DNS-molekulák; a citoplazmában különféle membránszervecskék találhatók.
A növényi sejteket vastag cellulóz sejtfal, plasztidok és egy nagy központi vakuólum jellemzi, amely a sejtmagot a perifériára tolja. A magasabb rendű növények sejtközpontja nem tartalmaz centriolokat. A raktározó szénhidrát a keményítő.
A gombasejtek kitint tartalmazó sejtmembránnal rendelkeznek, a citoplazmában központi vakuólum található, plasztidok nincsenek. Csak néhány gombánál van centriólum a sejtközpontban. A fő tartalék szénhidrát a glikogén.
Az állati sejteknek általában vékony sejtfaluk van, nem tartalmaznak plasztidokat és központi vakuolumot, a sejtközpontra a centriol jellemző. A tároló szénhidrát a glikogén.
Az eukarióta sejt felépítése
Egy tipikus eukarióta sejt három összetevőből áll: membránból, citoplazmából és sejtmagból.
Sejtfal
Kívül a sejtet egy héj veszi körül, melynek alapja a plazmamembrán, vagyis a plazmalemma, melynek jellegzetes szerkezete és vastagsága 7,5 nm.
A sejtmembrán fontos és nagyon sokrétű funkciókat lát el: meghatározza és fenntartja a sejt alakját; védi a sejtet a károsító biológiai ágensek behatolásának mechanikai hatásaitól; számos molekuláris jel (például hormonok) vételét végzi; korlátozza a sejt belső tartalmát; szabályozza a sejt és a környezet közötti anyagcserét, biztosítva az intracelluláris összetétel állandóságát; részt vesz az intercelluláris kontaktusok és a citoplazma különféle specifikus nyúlványainak (mikrovillák, csillók, flagellák) kialakításában.
Az állati sejtek membránjában található szénkomponenst glikokalixnek nevezik.
Az anyagcsere a sejt és környezete között folyamatosan zajlik. Az anyagok sejtbe és onnan történő szállításának mechanizmusa a szállított részecskék méretétől függ. A kis molekulákat és ionokat a sejt közvetlenül a membránon keresztül szállítja aktív és passzív transzport formájában.
Típustól és iránytól függően endocitózist és exocitózist különböztetünk meg.
A szilárd és nagy részecskék felszívódását és kiválasztását fagocitózisnak, illetve fordított fagocitózisnak, a folyékony vagy oldott részecskék - pinocitózisnak és fordított pinocitózisnak - nevezik.
Citoplazma
A citoplazma a sejt belső tartalma, és hialoplazmából és a benne elhelyezkedő különféle intracelluláris struktúrákból áll.
A hialoplazma (mátrix) szervetlen és szerves anyagok vizes oldata, amely megváltoztathatja a viszkozitását és állandó mozgásban van. A citoplazma mozgásának vagy áramlásának képességét ciklózisnak nevezik.
A mátrix egy aktív közeg, amelyben számos fizikai és kémiai folyamat játszódik le, és amely a sejt minden elemét egyetlen rendszerré egyesíti.
A sejt citoplazmatikus szerkezetét zárványok és organellumok képviselik. A zárványok viszonylag nem állandó jellegűek, bizonyos típusú sejtekben az élet bizonyos pillanataiban fordulnak elő, például tápanyag-utánpótlásként (keményítőszemcsék, fehérjék, glikogéncseppek) vagy a sejtből kiválasztandó termékek. Az organellumok a legtöbb sejt állandó és nélkülözhetetlen alkotóelemei, amelyek meghatározott szerkezettel rendelkeznek és létfontosságú funkciót látnak el.
Az eukarióta sejt membránszervecskéi közé tartozik az endoplazmatikus retikulum, a Golgi-készülék, a mitokondriumok, a lizoszómák és a plasztidok.
Endoplazmatikus retikulum. A citoplazma teljes belső zónája számos kis csatornával és üreggel van kitöltve, amelyek falai a plazmamembránhoz hasonló szerkezetű membránok. Ezek a csatornák elágaznak, összekapcsolódnak egymással és egy hálózatot alkotnak, amelyet endoplazmatikus retikulumnak neveznek.
Az endoplazmatikus retikulum szerkezetét tekintve heterogén. Két típusa ismert - szemcsés és sima. A szemcsés hálózat csatornáinak és üregeinek membránjain sok kis kerek test - riboszóma található, amelyek durva megjelenést kölcsönöznek a membránoknak. A sima endoplazmatikus retikulum membránjai nem hordoznak riboszómákat a felületükön.
Az endoplazmatikus retikulum számos különböző funkciót lát el. A szemcsés endoplazmatikus retikulum fő funkciója a fehérjeszintézisben való részvétel, amelyet a riboszómák hajtanak végre.
A sima endoplazmatikus retikulum membránjain lipidek és szénhidrátok szintetizálódnak. Mindezek a szintézistermékek csatornákban és üregekben halmozódnak fel, majd különféle sejtszervecskékbe kerülnek, ahol sejtzárványként elfogyasztják vagy felhalmozódnak a citoplazmában. Az endoplazmatikus retikulum köti össze a sejt fő organellumait.
golgi készülék
Számos állati sejtben, például az idegsejtekben, a sejtmag körül elhelyezkedő összetett hálózat formáját ölti. A növények és protozoonok sejtjeiben a Golgi-készüléket egyedi sarló- vagy rúd alakú testek képviselik. Ennek az organellumnak a szerkezete hasonló a növényi és állati szervezetek sejtjeiben, alakjának változatossága ellenére.
A Golgi-készülék összetétele a következőket tartalmazza: membránokkal határolt és csoportokban elhelyezkedő üregek (egyenként 5-10); az üregek végein található nagy és kis buborékok. Mindezek az elemek egyetlen komplexumot alkotnak.
A Golgi-készülék számos fontos funkciót lát el. Az endoplazmatikus retikulum csatornáin keresztül a sejt szintetikus tevékenységének termékei - fehérjék, szénhidrátok és zsírok - eljutnak hozzá. Mindezek az anyagok először felhalmozódnak, majd kis és nagy buborékok formájában bejutnak a citoplazmába, és vagy magában a sejtben hasznosulnak élettevékenysége során, vagy eltávolítják onnan és felhasználják a szervezetben. Például az emlősök hasnyálmirigyének sejtjeiben emésztőenzimek szintetizálódnak, amelyek az organoid üregeiben halmozódnak fel. Ezután enzimekkel teli hólyagok képződnek. A sejtekből a hasnyálmirigy-csatornába választódnak ki, ahonnan a bélüregbe áramlanak. Ennek az organoidnak egy másik fontos funkciója, hogy a membránján zsírok és szénhidrátok (poliszacharidok) szintetizálódnak, amelyeket a sejtben használnak fel, és amelyek a membránok részét képezik. A Golgi apparátus aktivitásának köszönhetően megtörténik a plazmamembrán megújulása és növekedése.
Mitokondriumok
A legtöbb állati és növényi sejt citoplazmája kis testeket (0,2-7 mikron) - mitokondriumokat (görögül "mitos" - fonal, "kondrium" - szemcse, szemcse) tartalmaz.
A mitokondriumok jól láthatóak egy fénymikroszkópban, amellyel láthatjuk alakjukat, elhelyezkedésüket, megszámolhatjuk a számukat. Belső szerkezet a mitokondriumokat elektronmikroszkóppal vizsgálták. A mitokondrium héja két membránból áll - külső és belső. A külső membrán sima, nem képez ráncokat, kinövéseket. A belső membrán éppen ellenkezőleg, számos redőt képez, amelyek a mitokondriumok üregébe irányulnak. A belső membrán redőit cristae-nak (lat. "crista" - fésű, kinövés) nevezik.A különböző sejtek mitokondriumaiban a cristae száma nem azonos. Több tíztől több százig lehet, és különösen sok a kriszta az aktívan működő sejtek mitokondriumában, például izomsejtekben.
A mitokondriumokat a sejtek "erőműveinek" nevezik, mivel fő funkciójuk az adenozin-trifoszfát (ATP) szintézise. Ez a sav minden élőlény sejtjének mitokondriumában szintetizálódik, és univerzális energiaforrás, amely szükséges a sejt és az egész szervezet létfontosságú folyamatainak végrehajtásához.
Az új mitokondriumok a sejtben már meglévő mitokondriumok osztódásával jönnek létre.
Lizoszómák
Kis kerek testek. Mindegyik lizoszómát membrán választja el a citoplazmától. A lizoszómán belül olyan enzimek találhatók, amelyek lebontják a fehérjéket, zsírokat, szénhidrátokat, nukleinsavakat.
A lizoszómák megközelítik a citoplazmába került táplálékrészecskét, összeolvadnak vele, és egy emésztési vakuólum keletkezik, melyben lizoszóma enzimekkel körülvett táplálékrészecske található. A táplálékrészecske emésztése során keletkező anyagok bejutnak a citoplazmába, és a sejt felhasználja őket.
A tápanyagok aktív emésztésének képességével rendelkező lizoszómák részt vesznek a létfontosságú tevékenység során elpusztuló sejtrészek, egész sejtek és szervek eltávolításában. A sejtben folyamatosan új lizoszómák képződnek. A lizoszómákban lévő enzimek, mint bármely más fehérje, a citoplazma riboszómáin szintetizálódnak. Ezután ezek az enzimek az endoplazmatikus retikulum csatornáin keresztül bejutnak a Golgi-készülékbe, amelynek üregeiben lizoszómák képződnek. Ebben a formában a lizoszómák belépnek a citoplazmába.
plasztidok
A plasztidák minden növényi sejt citoplazmájában megtalálhatók. Az állati sejtekben nincsenek plasztidok. A plasztidoknak három fő típusa van: zöld - kloroplasztiszok; piros, narancssárga és sárga - kromoplasztok; színtelen - leukoplasztok.
A legtöbb sejt számára kötelezőek azok az organellumok is, amelyeknek nincs membránszerkezetük. Ide tartoznak a riboszómák, mikrofilamentumok, mikrotubulusok és a sejtközpont.
Riboszómák. A riboszómák minden élőlény sejtjében megtalálhatók. Ezek 15-20 nm átmérőjű, lekerekített mikroszkopikus testek. Minden riboszóma két különböző méretű, kicsi és nagy részecskéből áll.
Egy sejt sok ezer riboszómát tartalmaz, ezek vagy a szemcsés endoplazmatikus retikulum membránjain helyezkednek el, vagy szabadon fekszenek a citoplazmában. A riboszómák fehérjékből és RNS-ből állnak. A riboszómák funkciója a fehérjeszintézis. A fehérjeszintézis egy összetett folyamat, amelyet nem egy riboszóma, hanem egy egész csoport hajt végre, beleértve akár több tucat kombinált riboszómát is. A riboszómák ezt a csoportját poliszómának nevezik. A szintetizált fehérjék először az endoplazmatikus retikulum csatornáiban és üregeiben halmozódnak fel, majd eljutnak az organellumokhoz és a sejthelyekhez, ahol elfogyasztják őket. Az endoplazmatikus retikulum és a membránjain található riboszómák egyetlen berendezés a fehérjék bioszintéziséhez és szállításához.
Mikrotubulusok és mikrofilamentumok
Különböző kontraktilis fehérjékből álló fonalas szerkezetek, amelyek a sejt motoros funkcióit okozzák. A mikrotubulusok üreges hengerek formájában vannak, amelyek falai fehérjékből - tubulinokból állnak. A mikrofilamentumok nagyon vékony, hosszú, fonalas szerkezetek, amelyek aktinból és miozinból állnak.
A mikrotubulusok és mikrofilamentumok behatolnak a sejt teljes citoplazmájába, kialakítva annak citoszkeletonját, ami ciklózist, az organellumok intracelluláris mozgását, a kromoszómák szétválását okozza az osztódás során. nukleáris anyag stb.
Sejtközpont (centroszóma). Az állati sejtekben a sejtmag közelében egy organoid található, amelyet sejtközpontnak neveznek. A sejtközpont fő része két kis testből áll - centriolokból, amelyek a sűrűsödött citoplazma kis területén helyezkednek el. Mindegyik centriole legfeljebb 1 µm hosszú henger alakú. A centriolok fontos szerepet játszanak a sejtosztódásban; részt vesznek a hasadási orsó kialakításában.
Az evolúció során a különböző sejtek alkalmazkodtak a különböző körülmények között élni és meghatározott funkciókat ellátni. Ehhez speciális organoidok jelenléte kellett bennük, amelyeket specializáltnak neveznek, ellentétben a fentebb tárgyalt általános célú organellumokkal. Ide tartoznak a protozoonok összehúzódó vakuólái, az izomrostok myofibrillumai, az idegsejtek neurofibrillumai és szinaptikus vezikulumai, a hámsejtek mikrobolyhjai, egyes protozoák csillói és flagellái.
Mag
A sejtmag az eukarióta sejtek legfontosabb alkotóeleme. A legtöbb sejtnek egyetlen magja van, de vannak többmagvú sejtek is (számos protozoonban, gerincesek vázizomzatában). Egyes nagyon specializált sejtek sejtmagokat veszítenek (például emlős vörösvértestek).
A mag általában gömbölyű vagy ovális alakú, ritkábban lehet szegmentált vagy fusiform. A mag a nukleáris membránból és a karioplazmából áll, amely kromatint (kromoszómákat) és magvakat tartalmaz.
A magburkot két membrán (külső és belső) alkotja, és számos pórust tartalmaz, amelyeken keresztül különböző anyagok cserélődnek a sejtmag és a citoplazma között.
A karioplazma (nukleoplazma) egy zselészerű oldat, amely különféle fehérjéket, nukleotidokat, ionokat, valamint kromoszómákat és sejtmagot tartalmaz.
A nucleolus egy kicsi, lekerekített test, intenzíven festődött, és nem osztódó sejtek magjában található. A nucleolus funkciója az rRNS szintézise és azok összekapcsolása a fehérjékkel, azaz. riboszóma alegységek összeállítása.
Kromatin - csomók, szemcsék és fonalas szerkezetek, amelyeket bizonyos színezékek specifikusan megfestenek, és amelyeket DNS-molekulák és fehérjék kombinációja képez. A kromatin összetételében lévő DNS-molekulák különböző részei eltérő fokú helicitást mutatnak, ezért különböznek a színintenzitásban és a genetikai aktivitás természetében. A kromatin a genetikai anyag létezésének egy formája a nem osztódó sejtekben, és lehetőséget ad a benne található információ megkettőzésére és megvalósítására. A sejtosztódás folyamatában a DNS spiralizálódik, és a kromatin struktúrák kromoszómákat alkotnak.
A kromoszómák sűrű, intenzíven festődő struktúrák, amelyek a genetikai anyag morfológiai szerveződésének egységei, és biztosítják annak pontos eloszlását a sejtosztódás során.
Az egyes biológiai fajok sejtjeiben a kromoszómák száma állandó. Általában a testsejtek magjában (szomatikus) a kromoszómák párban jelennek meg, az ivarsejtekben nem párosulnak. A csírasejtek egyetlen kromoszómakészletét haploidnak (n), a szomatikus sejtekben lévő kromoszómakészletet diploidnak (2n) nevezik. A különböző organizmusok kromoszómái mérete és alakja eltérő.
Kariotípusnak nevezzük a diploid kromoszómakészletet egy bizonyos típusú élő szervezet sejtjében, amelyet a kromoszómák száma, mérete és alakja jellemez. A szomatikus sejtek kromoszómakészletében a páros kromoszómákat homológnak, a különböző párokból származó kromoszómákat nem homológnak nevezzük. A homológ kromoszómák mérete, alakja, összetétele azonos (az egyik az anyai, a másik az apai szervezettől öröklődik). A kariotípusban lévő kromoszómák szintén fel vannak osztva autoszómákra, vagy nem nemi kromoszómákra, amelyek férfiak és nők esetében azonosak, valamint heterokromoszómákra, vagy nemi kromoszómákra, amelyek részt vesznek az ivar meghatározásában, és különböznek a férfiakban és a nőkben. Az emberi kariotípust 46 kromoszóma (23 pár) képviseli: 44 autoszóma és 2 nemi kromoszóma (a nősténynek két egyforma X kromoszómája van, a hímnek X és Y kromoszómája van).
A sejtmag tárolja és megvalósítja a genetikai információkat, szabályozza a fehérje bioszintézis folyamatát, és a fehérjéken keresztül - az összes többi életfolyamatot. A sejtmag részt vesz az örökletes információ leánysejtek közötti replikációjában és elosztásában, és ennek következtében a sejtosztódás és a szervezet fejlődésének szabályozásában.